Greener Journal of Agricultural Sciences
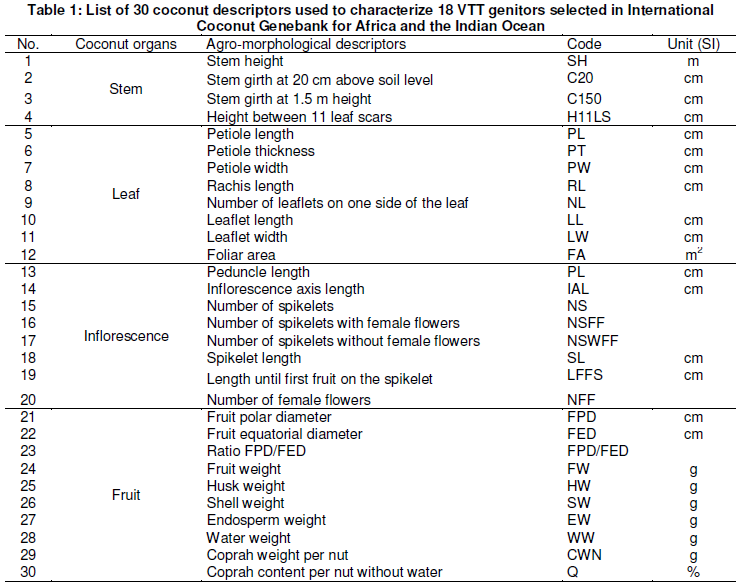
ISSN: 2276-7770; ICV: 6.15
Vol. 6 (4), pp. 134-144, April 2016
Copyright ©2017, the copyright of this article is retained by the author(s)
http://gjournals.org/GJAS

|
Greener Journal of Agricultural Sciences ISSN: 2276-7770; ICV: 6.15 Vol. 6 (4), pp. 134-144, April 2016 Copyright ©2017, the copyright of this article is retained by the author(s) http://gjournals.org/GJAS |
|
Research Article (DOI: http://doi.org/10.15580/GJAS.2016.4.020416032)
Morphological Diversity among 18 Genitors of Vanuatu Tall (VTT) Coconut (Cocos nucifera L.) Population used in Crosses for Hybrids Resistant to Lethal Yellowing Disease Selection at Port-Bouët, Côte d’Ivoire
Koffi Eric-BZ*1,2, Yao SDM3, Sie RS1, Konan KJL2,
Koffi Y1,2, Doh F1,2, Issali AE2, Allou K2
1UFR Sciences de la Nature, Nangui Abrogoua University, 02 BP 801 Abidjan 02, Côte d'Ivoire.
2Marc Delorme Research Station, CNRA, 07 BP 13 Abidjan 07, Côte d’Ivoire.
3UFR des Sciences Biologiques, Peleforo Gon Coulibaly University, BP 1328 Korhogo, Côte d’Ivoire.
|
ARTICLE INFO |
ABSTRACT |
|
|
Article No.: 020416032 DOI: 10.15580/GJAS.2016.4.020416032 |
Genetic control by creation of the coconut varieties expressing both Lethal Yellowing (LY) disease resistance and good agronomic performances led breeders to select in Tall populations which contain large diversity, some peculiar palms used as genitors. Thus, 18 genotypes of Vanuatu Tall (VTT) coded from G1 to G18 were described from 30 quantitative descriptors assessed on the coconut organs that are stem, leaf, inflorescence and fruit. The results showed that these 18 VTT genotypes were divided into two agro-morphological clusters: (i) genotypes providing large nuts with heavy components and (ii) the genotypes characterized by small fruit with weak component weights. Therefore, the genotypes that belong to first cluster G1, G4, G6, G7, G8, G9, G11, G13, G14, G16, G17 and specially G15 which also exhibited a vegetative robustness, could be selected as parent palms and involved in crosses with Dwarf or Tall coconut populations to develop hybrids that are potentially resistant to LY disease and producing large fruits with heavy components.
|
|
|
Submitted: 04/02/2016 Accepted: 25/01/2016 Published: 08/04/2016 |
||
|
*Corresponding Author Koffi Eric-BZ E-mail: zadjehi2003 @yahoo .fr Phone: (+225) 06000569 |
||
|
Keywords: Coconut palms, genitor VTT, genotype, morphological trait, Lethal Yellowing (LY) disease, selection, Côte d’Ivoire
|
||
|
|
|
ABBREVIATIONS
VTT: Vanuatu Tall; LY: Lethal Yellowing; MLOs: mycoplasma-like organisms; CNRA: Centre National de Recherche Agronomique; OPRI: Oil Palm Research Institute; ICG-AIO: International Coconut Genebank for Africa and the Indian Ocean; HS: Stem height; C20: Stem girth at 20 cm above soil level; C150: Stem girth at 1.5 m height; H11LS: Height between 11 leaf scars; PL: Petiole length; PT: Petiole thickness; PW: Petiole width; RL: Rachis length; NL: Number of leaflets on one side of the leaf; LL: Leaflet length; LW: Leaflet width; FA: Foliar area; PL: Peduncle length; IAL: Inflorescence axis length; NS: Number of spikelets; NSFF: Number of spikelets with female flowers; NSWFF: Number of spikelets without female flowers; SL: Spikelet length; LFFS: Length until first fruit on the spikelet; NFF: Number of female flowers; FPD: Fruit polar diameter; FED: Fruit equatorial diameter; FPD/FED: Ratio FPD/FED; FW: Fruit weight; HW: Husk weight; SW: Shell weight; EW: Endosperm weight; WW: Water weight; CWN: Coprah weight per nut; Q: Coprah content per nut without water;
INTRODUCTION
The coconut palm (Cocos nucifera L.) cultivated in the humid tropical regions is a perennial plant that belongs to the Arecaceae family. Two geographical origins of coconut palm were proposed by Gunn et al. (2011) which are Pacific and Indo-Atlantic oceanic basins. The coconut cultivation plays a key role in the economy of many countries where it has been nicknamed "tree of life" or "tree of hundred uses" (Bourdeix et al., 2005). Indeed, all parts of this palm species have many uses for human needs as a source of food, drink, fiber, construction material, charcoal and oil. However, in coconut palm cultivation, the fruit presents more economical and commercial interests because of its uses (Kanniah, 2005). Indeed, it is the oil extracted from mature kernel of the fruit that confers to the coconut palm its quality of oleaginous. Coconut oil is used in cooking, pharmaceuticals, industrial applications and biofuels (Gunn et al., 2011).
Nowadays, coconut palms cultivation around the world is threatened by Lethal Yellowing (LY) disease (Van Der Vossen and Chipungahelo, 2007). Already, LY disease has drastically affected and destroyed the coconut farms in many countries of the world such as the ones of East Africa (Tanzania, Kenya, Mozambique), West Africa (Nigeria, Togo, Ghana, Côte d’Ivoire), Caribbean (Jamaica, Haiti, Antilles) and Central America (Mexico, Honduras, Guatemala) according to Oropeza et al. (2005), Dollet et al. (2009) and Konan et al. (2013). Electron-microscope investigations found mycoplasma-like organisms (MLOs) responsible to LY disease in coconut palms (Rohde et al., 1993). Symptoms include leaf yellowing, starting in the oldest leaves and quickly move to the younger ones, drying to spikelet progressing to blackening of the whole inflorescence, rotting of heart, immature fruit drop and crown death of the palm after six months of initial symptoms appearance leaving a scenery of bare trunks as telephone poles (Konan et al., 2013). Today, an efficient chemical treatment to coconut LY disease management is not yet available. Therefore, all research works in Côte d’Ivoire and others countries are steered towards resistant varieties creation (Oropeza et al., 2005; Konan et al., 2013). Like that, the resistance tests were led since 1981 in the endemic zone of Ghana by the Centre National de Recherche Agronomique (CNRA) of Côte d’Ivoire in collaboration with Oil Palm Research Institute (OPRI) (Sangaré et al., 1992; Konan, 2005). At the end of these experimental trials, the coconut population of Vanuatu Tall (VTT) coming from the field genebank of Côte d’Ivoire was identified as the genetic sources controlling LY disease resistance. But, VTT have a bad agronomic performances (small fruit size, low fruit weight and coprah yield) (Bourdeix et al., 2005, Konan and Allou, 2006). To improve this coconut cultivar and vary LY disease resistance sources, CNRA decided to select the best individuals VTT to create together resistant hybrids with high yields. Also, to enhance knowledge of the agronomical potentials of coconut genitors implied in selection scheme, represents key information for improved seed production. The objective of this study was to know some morphological and agronomical characteristics of 18 best genotypes VTT selected as genitors for hybrids Tall x Tall or Dwarf x Tall resistant to Lethal Yellowing disease creation in coconut breeding programme of Côte d’Ivoire.
MATERIAL AND METHODS
Plant materials and measurements
Plant materials were constituted of 18 genotypes VTT coded from G1 to G18. Theses genotypes were planted at the density of 144 trees ha-1 in 1988 on plot No. 022 of the Marc Delorme station located at Port-Bouët Township between 5°14' N latitude and 3°55' W longitude. This station which belongs to Côte d’Ivoire, hosted the International Coconut Genebank for Africa and the Indian Ocean (ICG-AIO). Four climatic seasons with two rainy seasons (April to July and October to November) and two dry seasons (December to March and August to September) are known. From 1988 to 2010 at the meteorological station of Marc Delorme, the monthly average temperature ranged from 25.47 °C to 26.67 °C with 87.02% as mean value of relative humidity. Annual average of rainfall was 1742.37 mm. The soil of the Marc Delorme station is constituted of tertiary sands.
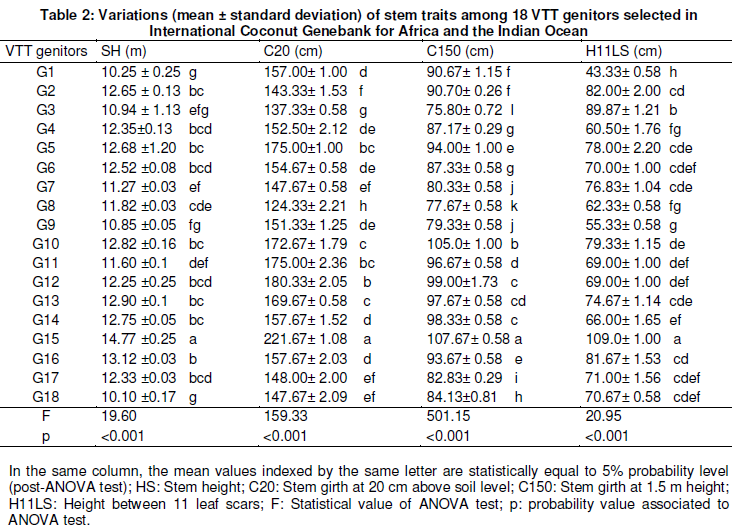
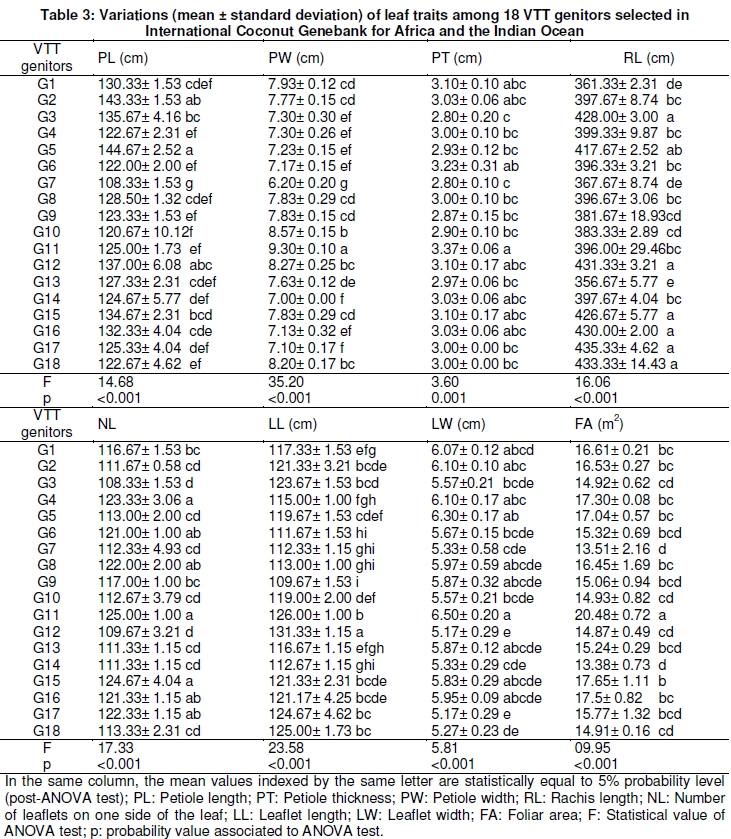
In the experiment, measurements were done on these VTT genitors in 2010 from 30 quantitative and agronomical traits including stem, leaf, inflorescence and fruits descriptors (Table 1) chosen in a long list of coconut descriptors were studied in detail by Yao et al. (2015).
Differences in measured traits between VTT genitors were tested by analysis of variance (ANOVA) incorporating mean comparison according to post-ANOVA test of Newman-Keuls at 5% threshold. The data were also subjected to hierarchical cluster analysis (HCA) based on Euclidean distances using an average fusion strategy UPGMA (Unweighted Pair-Group Method using Arithmetic average) (Baudouin and Santos, 2005) in order to estimate genetic relationship between 18 VTT genotypes. In addition, Student test was also used to reveal traits that mostly discriminated the clusters previously identified by HCA. All statistical analyses were performed with STATISTICA 7.1 (StatSoft France, 2005) software.
RESULTS
Variability of 30 agro-morphological traits studied in VTT genitors
The results from analysis of variance (ANOVA) showed that assessed agro-morphological traits discriminated significantly the 18 genitors VTT. Genotype G15 has the most values of stem height (SH = 14.77 m), stem girth at 20 cm above soil level (C20 = 221.67 cm) and stem girth at 1.5 m height (C150 = 107.67 cm) (Table 2). Low values of stem characteristics were observed in G1 (SH = 10.25 m), G18 (SH = 10.1 m), G8 (C20 = 124.33 cm) and G3 (C150 = 75.8 cm). G1 genotype was shown to have a low value of height between 11 leaf scars (H11LS = 43.33 cm) than the ones of other studied genotypes where the values ranged from 55.33 cm (G9) to 89.87 cm (G3).
Concerning leaf characteristics, the genotype G11 exhibited the most values of petiole width (PW = 9.3 cm), petiole thickness (PT = 3.37 cm) and foliar area (FA = 20.48 m2). The least values of petiole width (LGP = 7.1 cm), petiole thickness (PT = 2.8 cm) and foliar area (FA = 13.38 m2) were recorded in genotypes G17, G3 and G14 respectively (Table 3).
As for inflorescences characteristics, G18 genotype exhibited long peduncle (59.67 cm). G11 and G17 genotypes were produced along the inflorescence axis where mean values recorded varied from 42 to 43 cm (Table 4). But, inflorescence axis was less long in G6 (IAL = 28 cm), G1 (IAL = 30.33 cm), G7 (IAL = 28.67 cm) and G12 (IAL = 30 cm). G10 and G11 genotypes were produced per inflorescence and had the highest number of spikelets varying from 46 to 47 respectively. Low numbers of spikelets per inflorescence (NS = 31) were scored in G3, G6 and G12 genotypes. The highest values of spikelet length were detected in G15 (SL = 49.67 cm), G2 (SL = 47 cm), G13 (SL = 45.33 cm) and G14 (SL = 43.33 cm). Meanwhile, G6 and G9 genotypes were exhibited as the shortest length of spikelets where data varied from 31.33 to 40.67 cm. The first future fruit developed on the spikelet was more distant of the inflorescence axis in G15 genotype (LFFS = 11.33 cm) than the one observed in G12 (5.33 cm).
Measurements in fruit organ showed that all genotypes exhibited unrounded fruit shape with the ratio FDP/FED values superior to 1 (Table 5). Still, G11 and G12 genotypes were recorded higher values of ratio FPD/FED ranging from 1.67 to 1.73. The heaviest fruits were produced by G3 (FW = 837.33 g), G13 (FW = 908.33 g) and G15 (FW = 875 g) genotypes, meanwhile, low values of fruit weight were recorded in G2 (FW =503.33 g), G10 (FW = 525 g) and G18 (FW = 450 g) genotypes (Table 5). Husk weight in G15 (HW = 379.17 g) was superior to those of the other genotypes. The genotypes G15 (170.83 g), G13 (162.5 g), G16 (160 g) and G17 (153.33 g) were provided highest shell weight. High differences in coconut water weight distribution were observed among 18 VTT genitors (Table 5). Water weight variations were maximal in G13 and G17 with 150.83 g and 158.33 g respectively. Also, among studied genitors, endosperm weight variation was maximal in G4 (331.67 g) and minimum in G17 (163.33 g). G4 and G8 genotypes were given per nut higher yield of coprah with 176.02 g and 163.22 g respectively. Coprah contents per nut were ranged from 35.43 to 15.75% among 18 VTT genitors studied (Table 5).
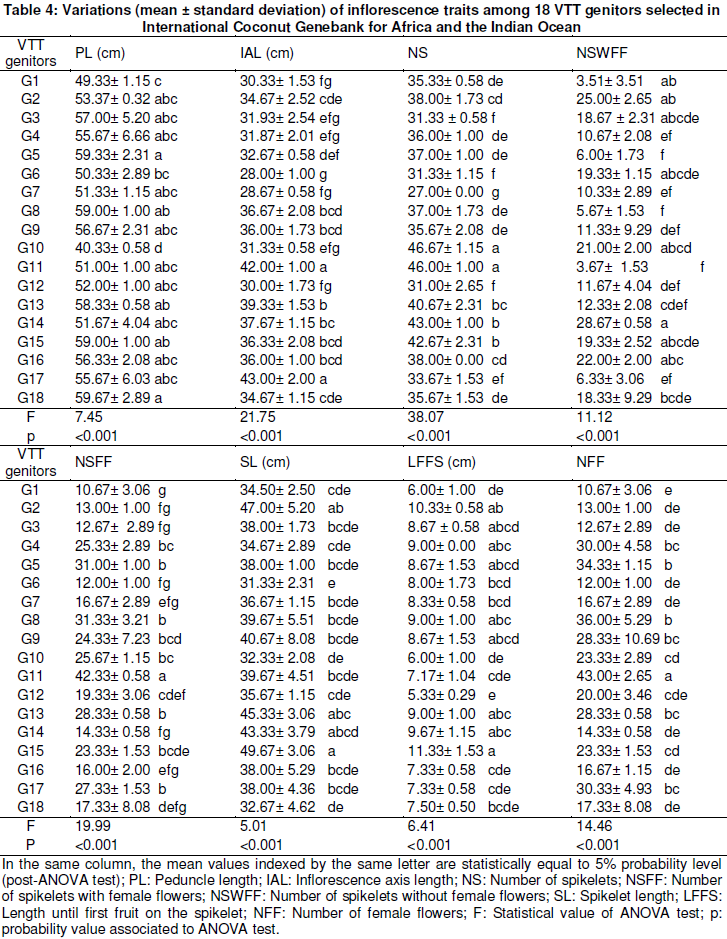
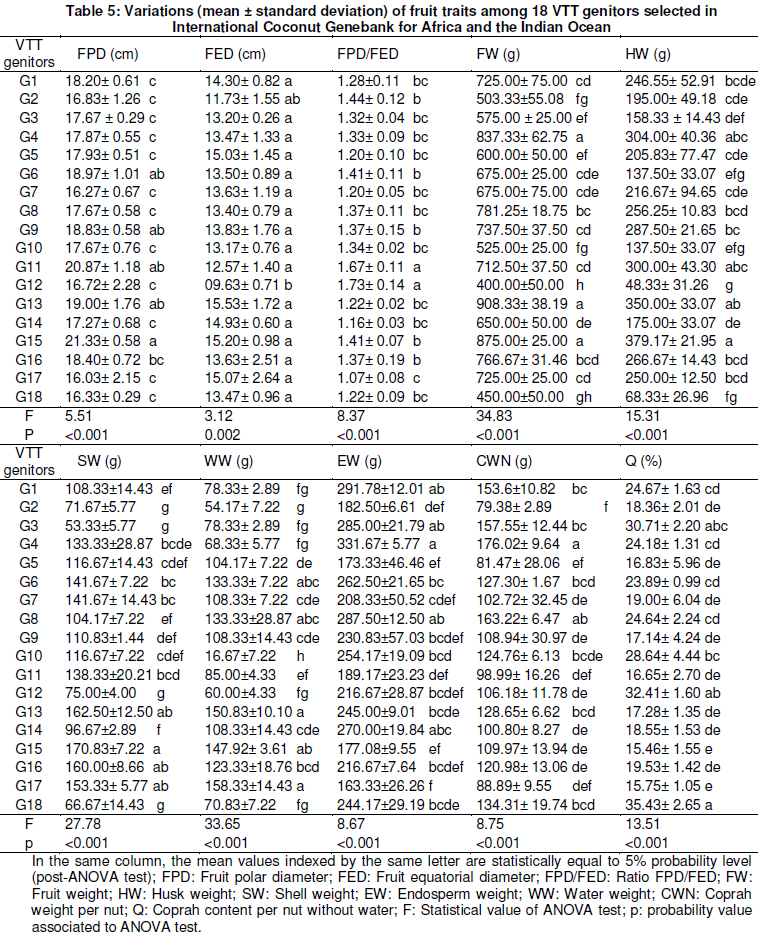
Agro-morphological pattern of 18 VTT genitors
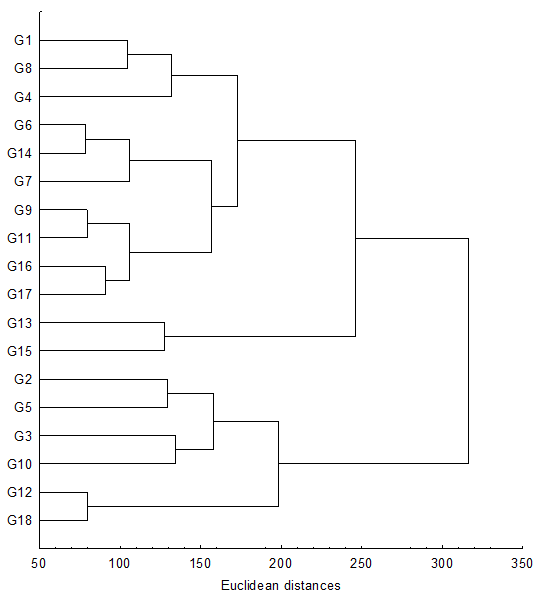
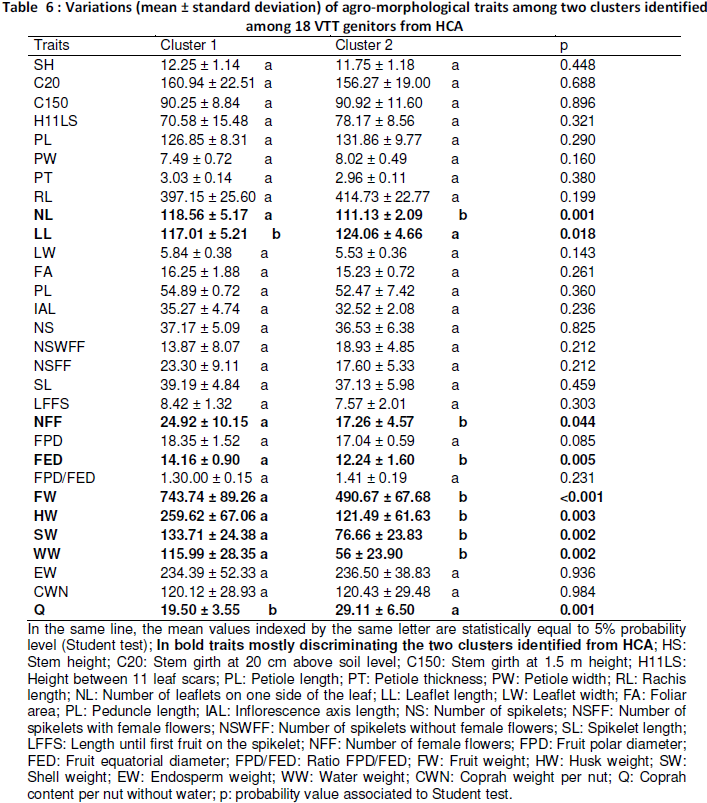
Hierarchical clustering analysis performed from UPGMA method divided 18 VTT genitors into two agro-morphological groups (Figure 1). The first cluster that was more varied was composed of the 12 genitors G1, G4, G6, G7, G8, G9, G11, G13, G14, G16, G17 and G15. The second cluster contains 6 genitors that are G2, G3, G5, G10, G12 and G18. These two clusters were significantly different from only 9 morphological traits where the ones of fruit morphology were more abundant (Table 6): fruit polar diameter (FPD), fruit weight (FW), husk weight (HW), shell weight (SW), coconut water weight (WW), coprah content per nut without water (Q), number of leaflets (NL), leaflet length (LL) and number of female flowers (NFF). Thus, mainly the genitors belonging to cluster 1 produced large size nuts with heavy components. But, the genitors of cluster 2 produced higher coprah content (Q = 29.11%) that those of the cluster 1 (Q = 19.5%). Furthermore, the genitors of the cluster 1 recorded more leaflets (NL = 119) on one side of the palm than the ones of cluster 2 (NL = 111). However, genitors of the cluster 2 exhibited higher leaflet length (LL = 123.33 cm). The genitors belonging to cluster 1 provided more female flowers (NFF = 25) that those of the second cluster (NFF = 17).
Figure 1: UPGMA dendrogram performed from 30 agro-morphological traits clustering 18 VTT selected in International Coconut Genebank for Africa and the Indian Ocean
DISCUSSION
This research was undertaken to study morphological and agronomical diversities among 18 genotypes coded from G1 to G18 selected in Vanuatu Tall (VTT) coconut population as genitors to create hybrid generations endowed genes controlling LY disease resistance and good agronomic performances. The results showed that G15 genotype develop very robust trunk. Thus, genitor G15 would have some genes that confer it the advantage to resist the violent winds and drought as reported by Konan et al. (2006). Concerning the high expression of foliar area, character G15 genotype occupied the second position after G11 genotype. So, on the contrary to the others genitors, G15 and G11 would have a high capacity to realize photosynthesis. Also, this high capacity to achieve photosynthesis could confer a stronger vegetative vigor, resistance to the diseases and pests in both G11 and G15 genotypes. It was mentioned that Tall coconut populations where these genitors VTT were chosen for the study are heterozygous and contain some resistance genes to the diseases and pathogens (Konan et al., 2006; Konan et al., 2007). Nevertheless, both low value of foliar area and higher leaf length were recorded in G3, G12, G16, G17 and G18 genitors. That could represent a handicap for the planting of these genitor descents. Indeed, the leaf length defines the density of plantation. This suggests that at the same plantation density as the ones of Tall x Tall hybrids (144 tree ha-1) and Dwarf x Tall hybrids (160 tree ha1), the progenies that come from genitor G15 which exhibited both higher foliar area and short leaf length, would express high yields of nut and coprah (Le Saint and De Nuce, 1987; Konan et al., 2010). G11 and G17 genotypes expressed higher length of inflorescence axis. This behavior could be an advantage for the progenies that come from these genitors. Indeed, the length of inflorescence axis influences the number and shape of the fruit (De Nuce and Wuidart, 1982). According to these authors, the longest inflorescences in coconut palm have more space to bear larger fruits. G11 and G12 genotypes express higher values of ratio FPD/FED. This suggests that these genotypes contain some genes determining more fruit numbers per unit of bunch than the others. Indeed, Bourdeix et al. (2005) reported that coconut populations such as VTT which produce high number of fruits per bunch have unrounded fruit shape (Bourdeix et al., 2005).
Hierarchical cluster analysis divided 18 VTT genitors into two agro-morphological groups. On one hand, there were the genitors producing high values of fruit size and fruit component weight (G1, G4, G6, G7, G8, G9, G11, G13, G14, G15, G16 and G17), on the other hand, the genitors were characterized by low weights of fruit and its components (G2, G3, G5, G10, G12 and G18). This reveals that fruit characters contribute mostly to coconut population discrimination as reported by Yao (2014). In relation to their discrimination, power fruit morphological characters have been used specifically as identification criteria of coconut varieties (Harries, 1981; Ashbumer et al., 1997). Our results indicate that stem, leaf and inflorescence traits could be consider as additional criteria for genetic diversity estimated within VTT coconut population. In previous review on coconut cultivars including Tall populations, Bourdeix et al. (2010) described three fruit sizes in VTT: large, medium and small. The large fruits have a weigh superior or equal to 900 g. The weight of the medium fruits ranges from 750 to 900 g and the smallest fruit is less than 750 g. Therefore following only fruit size, VTT characterized genotypes in our study are classified into the group of medium fruits for the genitors of the first cluster and smallest fruits for the genitors of the second cluster.
CONCLUSION
The present study aimed to assess 30 agro-morphological descriptors phenotypic diversity among 18 genotypes of Vanuatu Tall (VTT) involved in crosses Tall x Tall or Dwarf x Tall, to develop coconut hybrids expressing both LY disease resistance and high agronomic performance at Port-Bouët, Côte d’Ivoire. The results showed that in contrast to the others genitors studied G15 exhibited a very robust stem, high foliar area and short leaf length. From hierarchical cluster analysis, the 18 VTT genotypes were divided into two agro-morphological groups. Contrary to the second cluster (G2, G5, G3, G10, G12 and G18), the first cluster contains the genitors G1, G4, G6, G7, G8, G9, G11, G13, G14, G15, G16 and G17 which produce large fruits with heavy components. Based on the coconut hybridization programme of Côte d’Ivoire, genitors of the first cluster and especially genitor G15 which also exhibited a vegetative robustness may then be selected to generate hybrids expressing both LY resistance and large fruits with heavy components for replacement of susceptible coconut varieties.
REFERENCES
Baudouin L. and Santos G., (2005). Morphometric methodes of determining diversity in coconut. In Coconut Genetic Resources: Batugal P, Ramanatha Rao V, Olivier J (eds). International Plant Genetic Resources Institute: Selangor DarulEhsan, Malaisie; 209 – 224.
Bourdeix R., Konan J.L. and Labouisse J.P., (2010). Vanuatu Tall (VTT). In: Batugal P, Bourdeix R, Olivier JT, George MLC, editors. Catalogue of conserved coconut germplasm. International, Regional Office for Asia, the Pacific and Oceania, Serdang, Selangor Darul Ehsan, Malaysia: 292-293.
Bourdeix R., Konan J.L. and N’cho Y.P., (2005). Cocotier, guide des variétés traditionnelles et améliorées. CIRAD /CNRA ISBN 2-9525-4080-2, Montpellier (France), 58p.
De Nuce L. and Wuidart W., (1982). L’observation des caractéristiques de développement végétatif, de floraison et de production chez le cocotier. Oléagineux, 37 (6) : 291-300.
Dollet M., Jannot C., Baudouin L. and Cirad J.O., (2009). Le cocotier en Afrique et la maladie du jaunissement mortel. OCL, 16 (2): 74-75.
Gunn B.F., Baudouin L. and Olsen K.M., (2011). Independent Origins of Cultivated Coconut (Cocos nucifera L.) in the Old World Tropics. PLoS ONE 6(6): e21143, 8p.
Harries C., (1981). Identification pratique des variétés de cocotier. Oléagineux, 36 (2) : 62-79.
Kanniah J., (2005). Village-level production of high-value coconut products to enhance incomes of resource-poor coconut farmers. Coconut hybrids for smallholders. CFC / IPGRI, Amsterdam (Netherlands): 209-235.
Konan J.L. and Allou K.R., (2006). Jaunissement mortel du cocotier. Rapport de mission (Ghana 24-30 septembre 2006). Projet FSP, OPRI (Ghana)/CIRAD (France). 15p.
Konan J.L., (2005). Status of coconut genetic resources research in Côte d’Ivoire. Coconut genetic resources, edition Future Harvest ISBN 92-9043-629-8, IPGRI, Selangor Darul Ehsan (Malaisie): 654-660.
Konan J.L., Bourdeix R., Sangare A., Mondeil F., (2006). Caractérisation de quelques cultivars de cocotier (Cocos nucifera L.) tolérant à la sécheresse en Côte d’Ivoire. Agronomie Africaine 18 (2) : 145-156.
Konan J.L., Sie S.R., N’guetta S.P, Lekadou T.T. and Allou K., (2010). Assessment of vegetative growth and production of new improved coconut (Cocos nucifera L.) hybrids. Journal of Applied Biosciences 26: 1664-1674.
Konan J.L., Allou K., Atta-Diallo H., Saraka Yao D., Koua B., Kouassi N., Benabid R., Michelutti R., Scott J. and Arocha-Rosete Y., (2013). First report on the molecular identification of the phytoplasma associated with a lethal yellowing-type disease of coconut palms in Côte d’Ivoire. New Disease Reports 28, 3.
Konan K.J.N., Konan J.L., Koffi K.E., Lebrun P. and Sangare A., (2007). Coconut Microsatellite gene diversity analysis technology transfer to Côte d’Ivoire. Biotechnology 6 (3) : 383-388.
Le Saint J.P. and De Nuce L.M., (1987). Les hybrides de cocotiers nains : Performances et intérêt. Oléagineux, 42 (10) : 353-362.
Oropeza C., Escamilla J.A., Mora G., Zizumbo D. and Harrison N.A., (2005). Coconut lethal yellowing. Coconut genetic resources, edition Future Harvest ISBN 92-9043-629-8, IPGRI, Selangor Darul Ehsan (Malaisie) : 349-363.
Rohde W., Kullaya A., Mpunami A. and Becker D., (1993). Rapid and sensitive diagnosis of mycoplasma-like organisms associated with lethal disease of coconut palm by a specially primed polymerase chain reaction for the amplification of 16S rDNA. Oléagineux, 48 (7): 319-322.
Sangare A., Le Saint J.P., De Nuce L., (1988). Hybride de cocotiers prometteurs PB-122, PB-132, PB-214. Oléagineux, 43 (5) : 207-215.
Sangare A. De Taffin G., De Franqueville H., Arkhust E.D. and Pomier M., (1992). Le jaunissement mortel du cocotier au Ghana. Premier résultats sur le comportement au champ du matériel végétal. Oléagineux, 47 (12): 699-704.
StatSoft France, (2005). Statistica, logiciel d’analyse de données version 7.1. www.statsoft.fr.
Sugimura Y., Itano M., Salud C., Otsuji K. and Yamaguchi H., (1997). Biometric analysis on diversity of coconut palm: cultivar classification by botanical and agronomical traits. Euphytica 98: 29 -35.
Van Der Vossen, H.A.M. and Chipungahelo G.S.E., (2007). Cocos nucifera L. In: van der Vossen, H.A.M., Mkamilo, G.S. (Editeurs). PROTA 14 : Vegetable oils/Oléagineux. [CD-Rom]. PROTA, Wageningen, Pays Bas.
Yao S.D.M., (2014). Impact de la régénération de la collection internationale de cocotier (Cocos nucifera L.) pour l’Afrique et l’Océan indien sur la diversité agromorphologique et moléculaire. PhD thesis, Nangui Abrogoua University, Côte d’Ivoire ; 162 p.
Yao S.D.M., Konan J.L., Sie R., Diarrassouba N., Lekadou T.T., Koffi E.BZ., Yoboue K., Bourdeix R., Issali A.E., Doh F., Allou K. and Zoro Bi I.A., (2015). Fiabilité d’une liste minimale de descripteurs agromorphologiques recommandée par le COGENT dans l’étude de la diversité génétique du cocotier (Cocos nucifera L.). Journal of Animal and Plant Sciences, 26 (1): 4006-4022.
|
Cite this Article: Koffi Eric-BZ, Yao SDM, Sie RS, Konan KJL, Koffi Y1, Doh F, Issali AE, Allou K (2016). Morphological Diversity among 18 Genitors of Vanuatu Tall (VTT) Coconut (Cocos nucifera L.) Population used in Crosses for Hybrids Resistant to Lethal Yellowing Disease Selection at Port-Bouët, Côte d’Ivoire. Greener Journal of Agricultural Sciences, 6(4): 134-144, http://doi.org/10.15580/GJAS.2016.4.020416032. |