INTRODUCTION:
The germination of seed is said to follow a
sequential manner starting with seed imbibition which triggers resumption of
the metabolic activities therefore enforcing expression of the embryo and
emergence of the radicle (Miransari and Smith, 2009; Nonogaki et al.,
2010). Imbibition is a passive process and pressures
caused by swelling are not sufficient to cause a rupture of the surrounding
tissue. This is supported by the work of Mensah and Agbagwa (2004), who reported that embryo expansion is
repressed by ABA in some physiologically dormant seeds, so that imbibition
alone does not lead to complete seed germination. For complete germination,
process of imbibition is followed by activation of hydrolytic enzymes,
initiation of growth in the embryo, seed coat rupture and radicle emergence (Miransari and Smith, 2009). The different phases
of germination are required for offshoot of seedlings and hence to achieve
seedlings with quality yield and reduced disease attack; it is therefore
necessary to hasten germination rate for early radicle emergence (Singh et al., 2001; Subedi
and Ma, 2005).
Seeds have their different moisture requirements to achieve
germination known as critical seed moisture content and once that critical
level is achieved, the seed is then ready to initiate germination (Bewley et al., 2000). Finkelstein (2004)
also added that this action will cause an increase in the volume of the seed,
resulting to cracking of the testa which may differ
from emergence of seedling as seen in Brassicaceae and Solanaceae. Major events
after imbibition include DNA repair, initiation of respiration, mitochondrial
repair, restoration of cellular integrity, synthesis of germination-related
mRNA’s and protein. (Nonogaki et al., 2010). The DNA content in the radicle tip cells of wild-type tomato seeds was
reported to have increased prior to germination (Bino
et al., 1992). Bewley et al. (2013) reported that initiation
of DNA synthesis during germination is linked with DNA repair following
imbibition of dry seeds and also comes before cell division that follows
germination. Yanyan et al. (2018) further stated that the vigor of the seed can be
marked by the time of initiation of DNA replication, moreover it takes a longer
time for low quality seeds to achieve DNA repair prior to successful
replication.
This research focuses
on the germination and DNA studies of Cucumber (Cucumis sativus) seed treated with growth promoters.
MATERIALS AND
METHODS:
The matured
cucumber (Cucucmis sativus L.) seeds (Plate 1) were obtained from fruit
garden Port Harcourt, Nigeria. The seeds were properly identified by the
Curator at the Herbarium Unit of Department of Plant Science and Biotechnology,
University of Port Harcourt. Viability
test was carried out on the seeds to ascertain its viability; hence, non-viable
seeds were discarded. The viable seeds were surface-sterilized with ethanol for
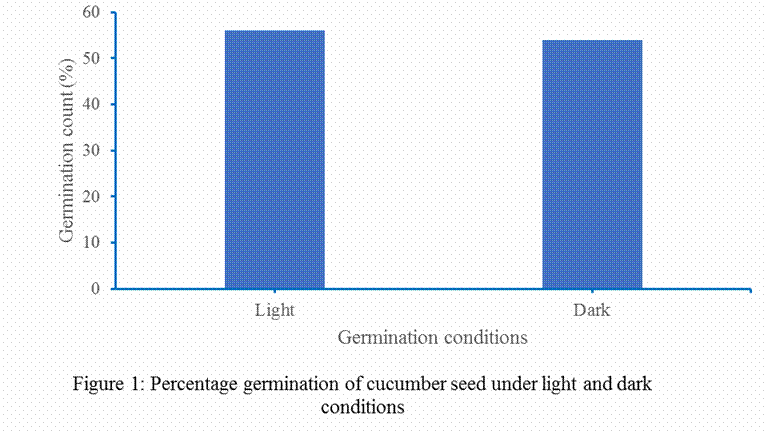
five (5) minutes and rinse with distilled water. Germination studies of cucumber
seeds were first carried out both under light and dark conditions.
The growth stimulants used in the study were gibberellic acid (GA3
– 350 g/mol), BAP (6-benzylaminopurine – 225.3 g/mol) and potassium
nitrate (KNO3 – 101.1 g/mol). The concentrations (1 mM, 5 mM and 10
mM) of these growth promoters were prepared, respectively. Water was used as
the Control treatment. These concentrations were used to pretreat cucumber
seeds with 20-seeds per batch. Each treatment was replicated five times. The
seeds were germinated under room temperature of 25oC, monitored
daily and the process lasted for 14 days. Germination
percentage of seeds taken for each treatment. Also, the DNA
concentration of the radicle with the highest germination count across
treatments were assessed using a Quick DNA miniprep kit for isolation of the
total DNA from the radicle sample of cucumber ensuring that there was no
contamination with RNA.
The data obtained from the study were subjected to statistical
analysis using SAS 9.1.3 version.
DISCUSSION:
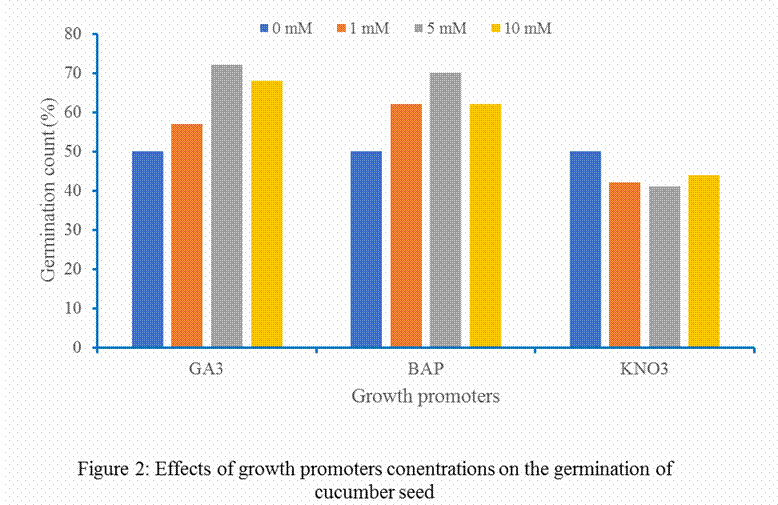
The findings
(Figure 2) indicated that GA3 and BAP enhanced percentage
germination compared to the Control. KNO3 treatments did not enhance
germination, as the germinations are comparable to Control treatment. The
effects of GA3 and kinetin in enhancing germination of dormant and
non-dormant seeds are well documented (Miyoshi and Sato, 1997; Mensah and Agbagwa, 2001; Zeb et al.,
2018) and supported the findings noted in this study.
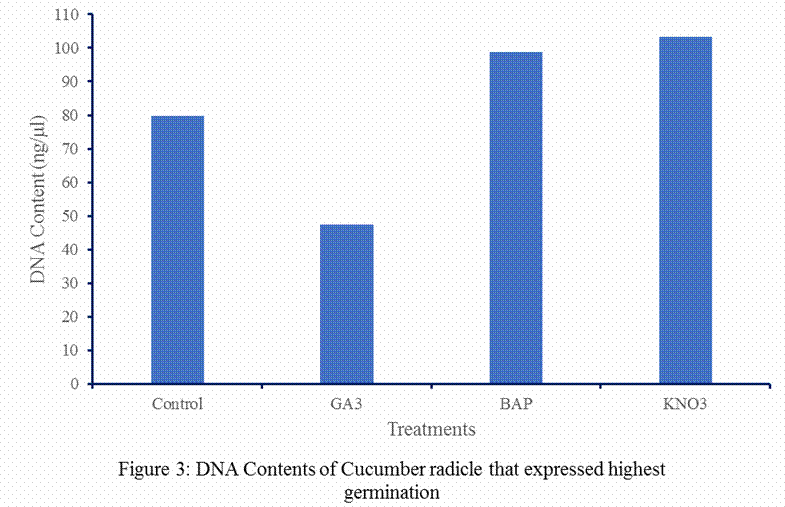
This study did not observe positive trend or relationship between
highest percentage germinations and DNA contents, indeed the reverse appeared
to be the case, that is, treatments with low germination gave high DNA contents
and those with high germinations, except 5 mM BAP that gave 70% germination and
recorded DNA content of 98.87 ng/µl. Gibberellin
and kinetin have been extensively reported to play a role in RNA and protein
synthesis, hydrolytic enzymes, substrate mobilization and elongation of embryo
axis in dormant and non-dormant seeds (Chrispeels and
Varner, 1967; Pinfield and Stobart,
1969; Jones and Armstrong, 1971; Varner and Ho, 1976; Jones
and Jacobsen, 1982; Vishal and Kumar, 2018). The study of the action of GA and kinetin has focused
on those molecular events that lead to de novo protein synthesis (Jones,
1973; Jacobsen et al., 1979; Jones and Jacobsen, 1982). The same
suggestions or arguments cannot be inferred in this study because treatments
(Control and KNO3) gave high DNA contents. However, it could be
suggested that GA and kinetin rather than act on the molecular level through de
novo synthesis, may act through the release of pre-formed enzymes
(hydrolases) in enhancing germination. It is suggested that further work need
to be undertaken to clearly establish the relationship between the hormone
enhancement of germinations and its molecular action.
The fact that GA3 and BAP effectively enhanced germination
is important for early emergence to avoid attack or damage the growing seeds
may encounter during unfavourable conditions (Singh et
al., 2001; Subedi and Ma, 2005). According to
Bewley et al. (2013), treated seeds initiate imbibition rapidly and
therefore hasten the phase II duration, making the interval between hydration
and radicle emergence short. Yanyan et al.
(2018) reported that the vigour of the seed can be
marked by the time of initiation of DNA replication, moreover it takes a longer
time for low quality seeds to achieve DNA repair prior to successful
replication. Gibberellic acid regulates the production of numerous enzymes by
activating the aleurone cells, notably alpha-amylase in growing cereals (Miransari and Smith, 2014).
CONCLUSION:
The study reveals that GA3, BAP
and KNO3 enhanced germination of cucumber seeds with 5 mM
concentration of GA3 and BAP been the concentration with highest
germination percentage while 10 mM for KNO3 treatment. Control
treatment had higher germination count than KNO3 treatment. The DNA
concentration of the cucumber radicle depends on the chemical used in treating
the seeds of cucumber and there is no clear relationship or trend between
treatments and DNA contents.
REFERENCES:
Bewley JD, Hempel FD, McCormick S and Zambryski P (2000). In: Buchanan BB, Gruissem W, Jones RL, (eds). Biochemistry
and Molecular Biology of Plants. American Society of Plant
Physiologists, Rockville, MD; Pp. 988–1403.
Bewley
JD, Kent J, Bradford K, Hilhorst H and Nonogaki H (2013). Seeds: Physiology
of Development, Germination and Dormancy, 3rd Edition. Springer New York;
Pp. 1-399.
Bino RJ, De-Vries JN, Kraak HL and Van-Pijlen JG
(1992). Flow cytometric determination of nuclear DNA
replication stages in tomato seeds during priming and germination. Annals
of Botany; 69: 231-236.
Chrispeels MJ and Varner JE (1967). Gibberellic acid-enhanced synthesis and release of
α-amylase and ribonuclease by isolated barley aleurone layers. Plant
Physiol., 42: 398-406.
Finkelstein
RR (2004). The role of hormones during seed development and
germination. In: Davies, P.J.
(Ed.), Plant Hormones: Biosynthesis,
Signal transduction, Action! The Netherlands, Kluwer Academic Publishers,
Dordrecht. Pp. 513–537.
Jacobsen JV,
Higgins TJV and Zwar JA (1979). Hormonal control of endosperm function during
germination. In The Plant Seed. eds. I. Rubenstein, B. G. Gegenbach,
R. L. Phillips and C. E. Green. Academic Press, New York, pp. 241-262.
Jones RL (1973). Gibberellins: their
physiological role. Ann. Rev. Plant Physiol. 24: 571-598.
Jones RL and
Armstrong JE (1971). Evidence for osmotic regulation of hydrolytic enzyme production in
germinating barley seeds. Plant Physiol., 48: 137-142.
Jones RL and
Jacobsen JV (1982). The role of endoplasmic reticulum in the synthesis and transport of
α-amylase in barley aleurone layers. Planta 156: 421-432.
Mensah
SI and Agbagwa IO (2001). The responses of seeds of Capsicum frutescens
L. to the exogenous application of some growth promoters. J. Agric. Biotechnol. Environ., 3:
37-47.
Mensah
SI and Agbagwa IO (2004). Breaking
dormancy in Gmelina arborea Roxb through treatment of seed with chemical substance and
alternating temperature. Journal of Biological
Research and Biotechnology; 2(1): 59 – 66.
Miransari
M and Smith DL (2009). Rhizobial lipochitooligossacharides and gibberellins enhance barley (Hordeum vulgare L.)
seed germination. African Journal of
Biotechnology, 8: 270-275.
Miyoshi K and Sato T (1997). The
effects of kinetin and gibberellin on the germination of dehusked
seeds of Indica and Japonica rice (Oryza sativa
L.) under anaerobic and aerobic conditions. Annals of Botany, 80:
479-483.
Nonogaki H, Bassel G and Bewly J (2010). Germination still a mystery. Plant Science, 179: 574-581.
Pinfield NJ and Stobart AK (1969). Gibberellic-stimulated nucleic and metabolism in
the cotyledons and embryonic axes of Coryllus avellane
(L.) seeds. New Phytol., 68: 993-999.
Singh
S, Singh P, Sanders DC and Wehner TC (2001). Germination
of watermelon seeds at low temperature. Cucurbit
Genetics Cooperative; 24: 59-64.
Subedi
KD and Ma BL (2005). Seed priming does not improve corn yield in
a humid temperate environment. Agronomy
Journal; 97: 211-218.
Varner JE and
Ho DT (1976). The
role of hormones in the integration of seedling growth. In The Molecular Biology of Hormone Action. ed. J. Papaconstantinou. Academic
Press, New York. pp. 173-194.
Vishal B and
Kumar PP (2018). Regulation of seed germination and abiotic stresses by gibberellins
and abscisic acid. Front. Plant Sci.,
9: 838
Yanyan
LV, Qing MO, Alison AP and Yanrong W (2018). DNA replication during seed germination, deterioration and its
relation to vigor in alfalfa and white clover. Crop Science; 58(3):
1393-1401.
Zeb A, Khan A, Khan R and Shabbir F (2018). Effect of different concentrations of kinetin on seed germination
in tomato. International Journal of Agronomy and Agricultural Research,
12(2): 1-8.