In Ethiopia onion covers an estimated annual
total production area of 22,767 ha both under rain-fed and irrigated condition
(CSA, 2015). However, onion production and productivity is constrained by
biotic and abiotic factors among which is soil salinity (Etissa
et al., 2014). According to Abebe et
al. (2015), out of 15,256.2 ha, Amibara irrigation scheme alone, 34% (5239.8 ha) of the
command area has been mapped as saline soil
electrical conductivity (EC) > 4 dSm-1 and sodium
absorption ratio (SAR) < 13). Expansion of the highly saline Basaka lake may aggravate salinity towards middle and lower
Awash basin, the East and Northeast direction, due to the topography of the
area (Olumana et al., 2009), where downstream irrigated
area production is highly constrained.
Salt accumulation can limit the
germination and development of various food crop species (Barroso
et al., 2010). This leads to morphological, cellular, biochemical and molecular
alterations that hinder the agricultural yield in response to the decrease in
the water potential of the soil solution, induced by the high osmolarity (Lima and Bull, 2008). In addition, ionic
toxicity promotes an imbalance in the absorption of essential nutrients,
causing metabolic disorders, which inhibit growth (Maia et al., 2012). Salt stress can also lead to
an excess intracellular production of reactive oxygen species (ROS) such as the
superoxide radical (O-2), the hydroxyl radical (OH-),
hydrogen peroxide (H2O2), and singlet oxygen (O2)
(Stanisavljevic et
al., 2011).
Salt stress may cause nutrient deficiencies
or imbalances, due to the competition of sodium (Na+) and chlorine (Cl–) with nutrients such as potassium (K+),
calcium (Ca2+) and nitrate (NO3-) (Hu
and Schmidhalter, 2005). It was observed that
increasing nitrogen (N) rates and salinity levels interacted to reduce chilli pod yield (Villa-Castorena
et al., 2003), while salt-stressed chilli performs well when adequately fertilized,
over-fertilization during early crop development may contribute to salinity and
decreased pod yield. It was also observed that foliar application of mono
potassium phosphate on onion crop was seriously affected at higher salinity of
irrigation water (4000 ppm), compared to moderate salinity level (2000 ppm)
(El-Dewiny et al.,
2013). In addition to increasing the salinity of irrigation water, it
caused a reduction in the contents of N, P, K, Ca and magnesium (Mg) nutrients, as a result of competition
between Na+, Cl- under high
saline water and these nutrients. Thus, a better understanding of the role of
mineral nutrients in plant resistance to drought and salinity will contribute
to an improved nutrient management in arid and semi-arid area such as Awash
Melkasa and in regions suffering from temporary drought.
Tolerance to salt stress is
complex physiological traits, metabolic pathway, and molecular or gene networks
(Gupta and Haung, 2014). However, adaptive response
to salt stress being identified varies within species and cultivars. Research
work in Turkey indicated that the effects of drought and salt tolerance of four
onion cultivars showed morphological and physiological variables differences
for both drought and salinity (Hanci and Cebeci, 2015). According to Beinsan
et al. (2015), great genetic diversity in terms of free proline
synthesis enables to identify cultivars that have a good tolerance to salt of
collected local land-races observed. In Ethiopia, there is scanty of
information regarding salt stress tolerance of onion cultivars. In view of this
the research was done with objectives of evaluating onion cultivars for yield
and yield components performance under salt stress levels on soil media.
MATERIALS AND METHODS
Description of the study site
The study was conducted at Melkasa Agricultural
Research Center (MARC) under open field from November 2017 to May 2018. MARC is
located 8°24’N latitude and 39°19’E with an altitude of 1550. The area receives
an average annual rainfall of 786 mm. the soil of the area is loam and clay
loam textural class. It has a dry climate with an average maximum and minimum
temperature of 35.4 and 20.63°C (MARC, 2017).
Experimental materials, treatments and
designs
Four released onion cultivars viz., Nafis, Nasic Red, Bombey Red and
Adama Red and five levels of root zone salinity (Awash water (0.3), 1.2, 4, 8
and 12 dSm-1) were factorially
combined; thus, 20 treatments (4 × 5) each replicated three times and the total
of 60 experimental units were laid down using factorial arrangements in
Randomized Complete Block Design.
Growing media preparation and field
management
Top soil from MARC field station and black
gravel were collected, sieved by mesh wire sized 2.36 mm and sterilized by
solar, separately. A composite made of 3 ratio top soil and 1 ratio sieved
gravel was mixed and one sample taken to MARC soil laboratory for soil physical
and chemical properties determination (Table 1). A total of 64 kg soil media
was filled in the box raised 40 cm from the and having 40 cm × 60 cm × 40 cm
volume with 2:1:3 layers, coarse gravels, sieved gravel alone and mix of sieved
gravel and top soil (3:1) from the bottom layers to upper layer, respectively
to facilitate easiness of drainage problems.
Uniform and vigor
seedlings were transplanted on January 24, 2018 by maintaining 5 and 10 cm
spacing between plants and rows, respectively having a total of 36 plants per
experimental unit sized 0.24 m2. Salinized irrigation water
application was started on February 12, 2018 or 19 days after transplanting
(DAT). It was applied using watering cane according to onion crop water
requirement calculated from climate data of MARC with the aid of CROPWAT 8.0
software and recently predetermined crop (Kc) of
onion at the center. The irrigation water was fixed to 0.8, 2.6, 5.2 and 7.6
dSm-1 by considering root zone salinity to be at 1.2, 4, 8 and 12
dSm-1 according to FAO (1985)’s (Equation 1) .
ECe= 1.5 ECw (1)
Where, ECw=
electrical conductivity of irrigation water, ECe =
Electrical conductivity at root zone. Leaching requirement calculation given by
FAO and SAFR (2002) (Equation 2) was used to maintain root zone soil salinity
to the designed levels. Thus, additionally 17% of Awash
water (0.3 dSm-1) was added to each experimental unit to maintain
designed salt level considering 90% leaching efficiency.
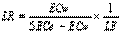 (2)
(2)
Where, LR= leaching requirement, ECw= electrical conductivity of irrigation water, ECe= electrical conductivity at root zone and LE= leaching
efficiency 90% used.
Pseudo-stem diameter
Non-significant differences were observed for
pseudo-stem diameter at 30 DAT for cultivars (P= 0.33), salt levels (P= 0.19)
as well as their interactions (P= 0.17). After two weeks of first stage (at 44
DAT), significant differences were observed among salt stress levels
(P=0.0001), while cultivars (P= 0.21) and their interaction (P= 0.33) did not
show significant differences. At 58 DAT, pseudo-stem diameter was highly
significantly influenced by salt stress levels (P=0.0001), whereas significance
differences were observed among cultivars (P=0.01). However, their interaction
did not show any significance (P= 0.36).
At the growth stage of 44 DAT, the highest
diameter was recorded with Awash water (0.3 dSm-1),
1.2 and 4 dSm-1 salt levels, and the thinnest value for 8 and 12 dSm-1.
As aging increased to 58 DAT, the highest stem diameter was also recorded with Awash water (6.30 mm), where the lowest value was recorded
with 8 and 12 dSm-1 salt stress level (Table 2). This result
indicated that increased salt stress decreased pseudo-stem diameter by bringing
physiological drought. This corroborates with the findings of Ghodke et al. (2018)
who reported that, increased drought stress in onion reduced pseudo-stem
diameter of the crop. Among the cultivars, Bombey Red showed the thickest
pseudo-stem diameter (5.36 mm) compared to Adama Red which had the thinnest
diameter. Our findings indicated that Bombey Red cultivar showed mild growth
performance at different stages of development in comparison to other
cultivars. This variation in performance was also observed under field
condition during the experiment compared to Adama Red which was dead at early
growth stage with the highest salt stress levels.
Leaf
length
Leaf length was highly and significantly
affected by cultivars (P= 0.01) and salt levels (P=0.0001), respectively.
However, leaf length has not been affected by their interaction (P=0.12). The
highest leaf length was recorded for Bombey Red, Nasic Red and Nafis while the
shortest plant height (19.13 cm) was recorded for Adama Red (Table 3). This
result confirms the field performance observed during the experiment where
Adama Red cultivar was very short and poorly performed. The main differences
seen among cultivars in plant height may be due to the intrinsic effects
existing among themselves.
Increase in salt
stress level decreased plant leaf height, where the highest leaf height 29.15
cm was recorded with Awash water (0.3) and 1.2 dSm-1
irrigated units and the shortest plant height was observed at 8 and 12 dSm-1
salt stress levels. Similar work was reported on onion leaf number and leaf
length that were negatively affected at high NaCl
concentrated irrigation water (Stab-Baba et al., 2010; Hanci
and Cebeci, 2015). Salt stress was reported to
inhibit plant height; increasing levels of NaCl in
the soil showed a diminished net photosynthetic rate, which automatically limited
the photosynthetic CO2 assimilation (Saleem
et al., 2011).
Leaf
width
ANOVA revealed significant differences among
cultivars (P=0.04) for leaf width and highly significant differences among salt
stress levels (P=0.001). However, their interaction did not show significant
differences (P= 0.24). The highest leaf width value was recorded for Nasic Red,
Bombey Red and Nafis while the lowest value (1.51 mm) was recorded for Adama
Red (Table 3). Our results indicated that Adama Red cultivar responded differently
from the rest cultivars via reducing its leaf size to the salt stress level. In
line with the report of Hernandez et al. (2003), salt stress inhibited cell
division and cell expansion; this consequently led to leaf expansion and leaf
width.
Of all the salt
stress levels, the widest leaf width (2.16 mm) was recorded with Awash water (0.3), whereas 1.2 and 4 dSm-1 showed
intermediate leaf width. The lowest value was recorded at 8 and 12 dSm-1
salt levels. Our results indicated that linear increase in concentration of NaCl stress levels significantly reduced leaf width of the
onion cultivars. Similar finding was also supported by the work of Munns (2002)
who reported that plants change their normal morphological structure in order
to defend themselves from stress. It was also reported that salinity reduced
final leaf width and emergency of number of lateral shoots in soybean. Dolatabadian et al. (2011) reported that salt reduced the
leaf area of beet root at the whole-plant level (leaf area ratio) and at the
individual leaf level (specific leaf area) (Rozema et
al., 2015).
Plant
height at full development
Analysis of variance showed that there were
significant differences among cultivars (P=0.01) and there were highly
significant differences observed among salt levels (P=0.001) in plant height.
Their interaction did not show any significance (P= 0.36). The tallest plant
height was recorded for Bombey Red and Nasic Red cultivars which were
statistically at par with Nafis, where the lowest plant height was recorded for
Adama Red (24.28 cm) cultivar.
Experimental units
irrigated with Awash water (0.3 dSm-1 and 1.2 dSm-1)
showed the highest plant height, whereas the lowest plant height was observed
at 8 and 12 dSm-1 salt levels (Table 3). At 4 dSm-1,
intermediate plant height was observed. Similar work was reported by Hanci and Cebeci (2015) in onion,
Girma et al. (2015) in rice; salinity concentration affected plant height
negatively. It was observed that onion plant height at highest salt levels (more than 4 dSm-1) gradually decreased and final
plants height was severely stunted. The plant height was stunted because enough
hormone was released to trigger leaf abscission (Dodd, 2005), and also due to
the earliest response of glycophytes to salt stress (Munns and Termaat, 1986). The
decrease in the availability of cytokinins may also
cause growth in inhibition of salt-stressed crops (Raghavendra,
1991). The observed reduction in the plant height may be considered as an
avoidance mechanism, which minimizes water loss by transpiration when the
stomata are closed (Acosta-Motos et al., 2017).
Furthermore, a decrease in leaf and pseudo-stem creates a reduction in all
aerial part sizes and in the plant height.
Shoot
fresh and dry weight
Shoot fresh and dry weights were
significantly and highly significantly affected by cultivars (P=0.04 and P=
0.0007); highly significant differences were observed among salt levels (P=
0.0001), respectively. Their interaction did not show any significant difference
(P= 0.22).
Nasic Red cultivar
showed the highest fresh weight (51.33 g) was statistically at par with Nafis,
whereas the lowest fresh weight was recorded for Adama Red (26.25 g). The
highest dry above ground biomass weight was also recorded for Nasic Red, Bombey
Red and Nafis cultivars, while the least dry weight value was recorded for
Adama Red (4.76 g) cultivar. The result showed that Adama Red cultivar was
relatively less performing not only in dry and fresh biomass weight, but also
in quantum yield, leaf number, leaf length, and plant height parameters. This
could be due to the variability in internal factors through which the crops
maintain their physiology and morphological characters from the induced stress.
The highest fresh
and dry biomass weight was recorded with Awash water
at 1.2 dSm-1 salt stress levels and the least were recorded at 8 and
12 dSm-1 salt stress levels. In this study, salt stress
significantly reduced plant fresh and dry weight of onion cultivars as shown
above (Table 3). These results are similar to those of tomato (Sholi, 2012), sunflower (Akram and Ashraf, 2011), mustard (Hayat et
al., 2011) and okra (Saleem et al., 2011; Azeem
et al., 2017). This reduction in biomass either in fresh or dry weight might be
due to salt stress that significantly reduced growth parameters and
photosynthetic attributes which finally reduced photo-assimilate production and
translocation (Azeem et al., 2017).
Chlorophyll
content
Statistical analysis revealed that
chlorophyll content measured by SPAD- meter showed highly significant (P=
0.001) differences for the main factors as well as their interaction at 49 DAT,
63 DAT and 77 DAT stages.
|
Table 4. Interaction effects of onion cultivars and levels of salt stress on
chlorophyll (in SPAD mmolm-2s- 1) and
stomatal conductance (in mmolm-2s-1 determined with a porometer) at three growth stages under MARC in 2018.

Physiological parameter
|
Cultivars
|
Salt levels
|
SPAD ( mmolm-2s- 1)
|
Porometer
(mmolm-2s-1)
|
|
49 DAT
|
63 DAT
|
77 DAT
|
51 DAT
|
65 DAT
|
79 DAT
|
|
Adama Red
|
0.3
1.2
4
|
17.76ab
8.19efg
20.36a
|
4.6gh
7.38cdefg
7.65cdefg
|
9.58bc
5.45ef
7.06cdef
|
135.35cd
178.41a
107.55defgh
|
105.32cd
100.94cd
109.19c
|
146.06a
83.81gh
87.55fgh
|
|
|
8
|
17.90ab
|
9.27bcd
|
4.88ef
|
76.19hij
|
90.4de
|
96.36defg
|
|
|
12
|
11.89cde
|
8.15cdef
|
4.24f
|
115.8cdef
|
76.92ef
|
125.37b
|
|
Bombey Red
|
0.3
1.2
4
|
8.62efg
7.64fg
18.103ab
|
6.03efg
9.82bc
9.27bcd
|
9.56bc
6.6cdef
14.5a
|
62.87j
106.07defgh
96.98efghi
|
99.54cd
89.01de
52.27g
|
108.28cd
89.78efgh
52.72j
|
|
|
8
|
8.98defg
|
2.65h
|
9.28bcd
|
82.93fghij
|
96.17cd
|
70.16i
|
|
|
12
|
8.6efg
|
8.79cde
|
9.53bc
|
74.49hij
|
60.79fg
|
99.22def
|
|
Nafis
|
0.3
1.2
4
|
7.07g
9.62defg
12.39cd
|
12.11b
29.88a
6.54defg
|
14.71a
6.5cdef
15.8a
|
129.44cde
107.43defgh
193.38a
|
66.01fg
62.65fg
144.10b
|
84.47gh
48.98j
102.11de
|
|
|
8
|
10.94def
|
5.79efg
|
6.97cdef
|
113.28cdefg
|
105.13cd
|
71.61i
|
|
|
12
|
6.257g
|
7.25defg
|
5.96def
|
69.62ij
|
98.07cd
|
79.07hi
|
|
Nasic Red
|
0.3
1.2
4
|
14.823bc
7.81fg
9.87defg
|
9.53bcd
9.93bc
4.91gh
|
11.2b
8.3bcde
6.79cdef
|
168.11ab
81.02ghij
143.06bc
|
133.16b 187.5a
147.50b
|
144.38a
114.81bc
96.04defg
|
|
|
8
|
8.97defg
|
5.36fgh
|
5.54ef
|
139.58bcd
|
71.27f
|
89.22fgh
|
|
|
12
|
17.79ab
|
10.46bc
|
5.917def
|
129.31cde
|
95.43cd
|
84.64gh
|
|
|
CR (5%)
|
3.219
|
2.77
|
2.99
|
29.57
|
15.77
|
11.42
|
|
|
CV%
|
16.56
|
19.32
|
22.03
|
15.64
|
9.68
|
7.52
|
|
|
|
|
|
|
|
|
|
Means in the
column followed by the same letter(s) are not significantly different at 5%
level of significance. CR (0.05) = Critical Range at the 5 % level; and CV
(%) = coefficient of variation in percent.
|
The interaction
effects indicated that the highest SPAD value was recorded for Adama Red at 4
dSm-1 (20.36 mmolm-2s-1),
it was statistically at par with Adama Red with Awash water and 8 dSm-1
and also Nasic Red at 8 dSm-1. Nafis with 1.2 dSm-1 (29.88 mmolm-2s-1) at second stage and
Nafis with Awash water and 4 dSm-1, Bombey Red with 4 dSm-1
showed the highest SPAD values at third stage (Table 4).
The least SPAD value interacted with Nafis at 12 dSm-1 (6.25 mmolm-2s-1), Adama Red with
Awash water (4.6 mmolm-2s-1) and Adama Red at 12 dSm-1
salt levels (4.24 mmolm-2s-1), at 49 DAT, 63 DAT and 77
DAT, respectively. Thus, the result implied that each cultivar had independent
response to maintain water content of leaves and leaf chlorophyll after
prolonged stress duration may indicate a potential mechanism of osmotic
adjustment in low to moderately high salinity (Stavridou
et al., 2017). However, it was reported that SPAD value of chlorophyll
decreased significantly in the stressed leaves, because of salinity which
either inhibits synthesis and/or accelerates the degradation of the existing
chlorophyll molecules (Wani et al., 2013).
Stomatal conductance
ANOVA for the main factors and their
interaction effects on stomatal conductance (gs) determined by porometer
across the three stages (51 DAT, 65 DAT, and 79 DAT) showed highly significant
variances (P= 0.001). The highest gs was recorded for Nafis at 4 dSm-1 and Adama Red
at 1.2 dSm-1 which was statistically at par with Nasic Red with
Awash water at first stage. Nasic Red with 1.2 dSm-1 (187.5 mmolm-2s-1)
at second stage and Awash water with Adama Red and
Nasic Red showed the highest gs at third stage (Table
4). The least gs was observed in Bombey Red with
Awash water (62.87 mmolm-2s-1) at 51 DAT, which was not
statistically different from Adama Red with 8, Bombey Red with 8 and 12, Nafis with 12 and Nasic Red with
1.2 dSm-1. At 65 DAT, Bombey Red with 4
dSm-1 (52.27 mmolm-2s-1)
which was par with Bombey Red with 12, Nafis with Awash water and 1.2 dSm-1
and at third stage Bombey Red with 4 dSm-1 and Nafis with 12 dSm-1
showed the least gs. Likely Azeem
et al. (2017) reported that increased salt levels decreased gs
of okra cultivars. A significant decrease in the gs
of plants exposed to increased levels of salt stress diminished net
photosynthetic rate, by limiting internal CO2 concentration and
transpiration rate (Saleem et al., 2011). Although
the current study did not show linear increase or decrease of gs on salt stress levels because cultivars responded
independently, salt stress levels above 8 dSm-1
were highly affected leaf gs.
Bulb length and width
Two ways of analysis showed highly significant
variations for bulb length and bulb width among the cultivars (P=0.004). The
analysis also depicted highly significant differences of bulb length and width
among the salt levels (P= 0.0001). Their interaction did not show any
significant differences for bulb length and width (P= 0.19).
The highest bulb length was recorded for
Bombey Red and Nasic Red, whereas the highest bulb width was recorded for
Nafis, Bombey Red and Nasic Red. The least bulb length (26.39 mm) and width
(15.18 mm) were recorded for Adama Red cultivar. Although, it is difficult to
definitely determine performance based on only bulb length and width, due to
its dependence on nature of crop bulbs shape. This result indicated that Adama
Red cultivar was reduced in bulb length and width than others. Table 6
indicated that for Nafis cultivar, bulb length showed medium length; while bulb
width was the highest due to its nature of bulb shapes. Generally, the result
indicated that Bombey Red, Nasic Red and Nafis were the most performing cultivars
in bulb width.
The highest bulb length and width were
recorded on Awash water (0.30 dSm-1) and
1.2 dSm-1 salt levels; whereas the least bulb length and width were
observed at 8 and 12 dSm-1 salt levels. This result showed that an
increased salt concentration levels more than threshold (1.2 dSm-1)
radically reduced bulb length and width. Supportive work was reported by Kahouli et al. (2014)
in carrot; increase in salt concentration decreased root diameter and length.
The reduction in bulb length and width under high NaCl
salt concentration could be because salts induced internal water deficit which
caused partial or complete closure of stomata. Azeem
et al. (2017) finally inhibited leaf
expansion, reduced net photosynthetic capacity of the plants, leading to
reduction in biomass production (Saleem et al.,
2011).
Bulb
fresh and dry weight
Fresh and dry bulb biomass weights were significantly
influenced by cultivars (P=0.02 and P=0.03), respectively. There were also
highly significant differences in fresh and dry bulb biomass weights observed
among salt levels (P= 0.0001). However, their interaction did not show any
significance (P=0.46) for the two parameters.
The highest fresh and dry bulb biomass
weights were recorded for Bombey Red and Nasic Red which were statistically
similar with Nafis; whereas the least fresh and dry bulb biomass weight was
recorded for Adama Red (28.15 g) (Table 6). The result pinpointed Bombey Red,
Nasic Red and Nafis cultivars to perform better than Adama Red cultivar, which
has relatively low performance. The differences among cultivars might be due to
internal factor by which they maintain their morphological and physiological
parameters. This finally influences their dry matter accumulation and yield.
The highest fresh bulb biomass weight was
recorded on Awash water (0.3 dSm-1) and 1.2
dSm-1, whereas the least was recorded on 8 and 12 dSm-1
salt stress levels. However, the least dry bulb biomass weight was recorded on
12 dSm-1 which was statistically similar with 8 dSm-1;
whereas the highest dry biomass was recorded on Awash
water and 1.2 dSm-1 which were statistically at par at 4 dSm-1
salt level. The result indicated that increase in salt concentration reduces
fresh and dry biomass of onion cultivars. It was also reported by Backhausen et al.
(2005) that fresh and dry weight decreased by 30% due to the increase in NaCl more than 5 dSm-1.
There was an increase in dry weight at 1.2 dSm-1 irrigated units
than the control. This might be because salt concentration to certain limit may
increase total soluble solid. Hepksoy (2004) reported
that salinity increases total sugar contents and all sugar fractions of fruits Sastuma madrin c. Owari. Up to 4 dSm-1 salt levels of irrigation
water mild dry weight and fresh biomass weights were recorded.
Total
soluble solid (TSS)
TSS was significantly and highly affected by
cultivar (P=0.04) and salt levels (P= 0.0002), respectively. Statistical
analysis did not reveal significant differences among their interaction p=
0.30.
The highest TSS was recorded for Nasic Red (12.47 obrix),
while the lowest Brix was observed in Bombey Red, Adama Red and Nafis
cultivars. Under natural condition Nasic Red and Nafis are known to have the
highest 10- 18 oBrix of TSS while Bombey
Red was the least of all (Zelleke and Derso, 2015). Under current study similar result was found
in respective of their proportion. This variation in TSS among the cultivars
might be due to their genetic constituents.
The highest TSS in 12.78 oBrix
was observed on 1.2 dSm-1 which was statistically similar with 4 dSm-1
salt stress level; whereas the least was recorded on
12 dSm-1 (10.94 oBrix) which
was statistically at par with 0.3 dSm-1 and 8 dSm-1 treated
units (Table 6). This indicated that salt stress to certain level might
increase TSS as compared to the lowest salt concentration. Similar work was
reported by Ghodke et al. (2018) that under forced drought stress TSS was
slightly elevated in comparison to routinely irrigated plot. Hepaksoy (2004) also reported that Sastuma madrin c. Owari
orange orchard grown near sea compared to that farthest from the sea showed the
highest TSS. The increase in these TSS was reported to regulate its osmosis,
and improve its metabolic processes during stress conditions (Ripoll et al., 2014).
CONCLUSIONS
Salt stress is limiting onion production and
productivity due to crop sensitivity to salt stresses. Our study results
indicated that growing onion cultivars with salt stressed water significantly
affected growth variables, physiology, yield and yield components of onion
across the growing stages. Onions leaf length, leaf width, fresh and dry above
ground biomass weight, fresh and dry bulb biomass weights, TSS, bulb length and
width were affected significantly (p≤0.05) by cultivars and highly significantly
(p≤0.001) by salt levels. However, during early growth stage of up to 4 dSm-1 did not affect the number of
leaves per plant, pseudo stem diameter, plant height, but gradually decreased
after 58 DAT. The highest number of leaves per plant, plant height and pseudo
stem diameters were observed in Bombey Red and Nasic Red, which were
statistically similar with Nafis; whereas Adama Red cultivar showed the lowest
number of leaves, shortest and thinnest across the stages. Physiological
parameters like chlorophyll and stomatal conductance taken across the stages
showed highly significant variations among the main factors and their
interactions; whereas quantum yield of PSII showed significance variation at 40th
DAT and 68th DATs among cultivars and salt levels at 40 and 54 DATs
stages. The highest quantum yield was recorded for Nasic Red, Nafis and Bombey
Red, where Adama Red showed the least. Quantum yield was reduced as salt
concentration level increased. Bombey
Red, Nafis and Nasic Red cultivars showed the highest performance in leaf
length and width, plant height, bulb length and width, fresh and dry bulb
weight, dry above ground biomass. The highest leaf length, plant height, leaf
width, TSS (12.78 oBrix), dry bulb weight
(11.32 g), fresh and dry above ground biomass weight (70.01 g and 11.67g), bulb
length and width were recorded at 1.2 dSm-1 salt stress level.
Generally, most of the variables during early growth stages were not affected up to 4 dSm-1; whereas the highest growth and
yield performances were recorded at 1.2 dSm-1 salt stress levels. It
is concluded that our cultivars cannot resist salt stress beyond 4 dSm-1 and
Bombey Red, Nafis and Nasic Red can be used for salt levels less than 4 dS/m. However, the experiment should be repeated under
controlled environment adding more cultivars and less than 4 dSm-1 salt
levels in the future.
CONFLICT
OF INTERESTS
The authors have not declared any conflict of
interests.
ACKNOWLEDGEMENTS
The authors thank Ethiopian Institute of
Agricultural Research for financial support. They also thank Melkasa
Agricultural Research Center for their contribution to facilitate the working
environment.
REFERENCES
Abebe TF, Alamirew T, Abegaz F (2015). Appraisal And
Mapping of Soil Salinity Problem in Amibara Irrigation
Farms. Middle Awash Basin, Ethiopia.
Acosta-Motos
R, Ortuno MF, Bernal-Vicente A, Diaz-Vivancos P, Sanchez-Blanco MJ, Hernandez JA (2017). Plant
Responses to Salt Stress: Adaptive Mechanisms. Agronomy 7(1):18.
Akram NA, Ashraf M (2011). Pattern
of Accumulation of Inorganic Elements in Sunflower (Helianthus annuus L.) Plants Subjected to Salt Stress and
Exogenous Application of 5-Aminolevulinic Acid. Pakistan Journal of Botany
43(1):521-530.
Azeem A, Wu Y, Xing D, Javed
Q, Ullah I (2017). Photosynthetic
Response of Two Okra Cultivars under Salt Stress and Re-Watering.
Journal of Plant Interactions 12(1):67-77.
Backhausen JE, Klein M, Klock
M, Jung S, Scheibe R (2005). Salt
Tolerance of Potato (Solanum tuberosum
L. Var. Desiree) Plants Depends on Light Intensity and Air Humidity.
Plant Science 169(1):229-237.
Barroso CM, Franke LB, Barroso IB (2010). Substrate Luznagerminacao Das Sementes De Rainha-Do-Abismo. Horticultura Brasileira
28:236-240.
Beinsan C, Sumalan R, Vatca S (2015). Influence of Salt Stress on Quality of Some
Onion (Allium cepa L.) Local Landraces. Bulletin USAMV, Series Agriculture 72:2.
Central
Statistical Agency (CSA) (2015). Agricultural Sample Survey Volume III, Report On-Farm Management
Practices. Statistical Bulletin 578.
Chang PT (2003). Changes of flavor components of onion (Allium cepa L.) in a saline environment (Doctoral dissertation, uga). The University of Georgia.
Cosgrove D (1986). Biophysical Control of Plant Cell Growth. Annual Review of
Plant Physiology 37(1):377-405.
Dodd IC (2005). Root-to-Shoot
Signaling: Assessing the Roles of up in the up and down World of Long-Distance
Signaling in Planta. Plant and Soil 274(1-2):251-270.
Dolatabadian A, Sanavy S, Ghanati F (2011). Effect of Salinity on
Growth, Xylem Structure and Anatomical Characteristics of Soybean. Notulae Scientia Biologicae 3(1):41-45.
El-Dewiny CY, Hussein MM, Awad F
(2013). Influence of Mono
Potassium Phosphate Fertilizer on Mitigate the Negative Effects of High Saline
Irrigation Water on Onion. Crop Middle East Journal of Agriculture Research
2(4):152-158.
Etissa E, Dechassa N, Alamirew T, Alemayehu Y, Desalegn L (2014). Irrigation Water Management Practices in
Smallholder Vegetable Crops Production: The Case of the Central Rift Valley of
Ethiopia Science,
Technology and Arts Research Journal 3(1):74-83. FAO (1985). Water Quality for Agriculture. 186. In Ayers R.S. and Westcot D.W. eds. Irrigation and Drainage Paper 29:1.
Food and Agriculture Organization of the United Nations
(FAO), Sub-Regional Office for East and Southern Africa (SAFR) (2002). Irrigation Manual Planning, Development Monitoring and Evaluation
of Irrigated Agriculture with Farmer Participation, Eds Andreas P. Savva.
Karen Frenken.
V. I. Mod 1 - 6. Harare.
Ghodke PH, Andhale
PS, Gijare UM, Thangasamy
A, Khade YP, Mahajan V,
Singh M (2018).
Physiological and Biochemical Responses in Onion Crop to Drought Stress.
International Journal of Current Microbiology and Applied Sciences
7(1):2054-2062.
Girma BT, Ali HM, Gebeyaneh AA (2015). Effect of Salinity
on Final Growth Stage of Different Rice (Oryza
sativa L.) Genotypes. Asian
Journal of Agricultural Research 11:1- 9.
Gupta B, Huang B (2014).
Mechanism of Salinity Tolerance in Plant: Physiological, Biochemical and
Molecular Characterization. International Journal of
Genomics. Article ID 701596.
Hanci F, Cebeci E
(2015). Comparison of Salinity and Drought Stress Effects on
Some Morphological and Physiological Parameters in Onion (Allium Cepa L.) during Early Growth Phase. Bulgarian Journal
of Agricultural Science 21(6):1204-1210.
Hayat S, Mir BA, Wani AS, Hasan SA, Irfan M, Ahmad A (2011). Screening of
Salt-Tolerant Genotypes of Brassica juncea based on Photosynthetic Attributes.
Journal of Plant Interactions 6(1):53-60.
Hepaksoy S (2004). Effect of
Salinity on Some Fruit Quality Attributes and Sugar Composition of Satsuma mandarin cv. Owari.
Asian Journal of Plant Sciences 3(6):660-665.
Hernandez JA, Aguilar AB,
Portillo B, Lopez-Gomez E, Beneyto JM, García-Legaz MF (2003). The Effect of
Calcium on the Antioxidant Enzymes from Salt-Treated Loquat and Anger Plants.
Functional Plant Biology 30(11):1127-1137.
Hu Y, Schmidhalter
U (2005). Drought and Salinity: A Comparison of Their Effects on Mineral
Nutrition of Plants. Journal of Plant Nutrition and Soil Science
168(4):541-549. IPGRI, ECP/GR, AVRDC (2002). Descriptors for Allium (Allium
spp.).
International Plant Genetic
Resources Institute, Rome, Italy; European Cooperative Programme for Crop
Genetic Resources Networks (ECP/GR), Asian Vegetable Research and Development
Center, Taiwan
Kahouli B, Borgi Z, Hannachi C (2014). Study of the Tolerance of Ten Accessions
of Carrot (Daucus carota L.) to
Salinity. Journal of Stress Physiology and Biochemistry 10(3):18-27.
Lima MD, Büll
LT (2008). Produção de cebola em
solo salinizado. Revista Brasileira
de Engenharia Agrícola e Ambiental, pp. 231-235.
Machado RMA, Serralheiro RP (2017). Soil Salinity: Effect on Vegetable Crop Growth. Management
Practices to Prevent and Mitigate Soil Salinization. Horticulture
3(2):30.
Maia JM, Ferreira-Silva SL,
Voigt EL, Macedo CD, Ponte LFA, Silveira
JAG (2012). Atividade De Enzimas Antioxidantes
E Inibição Do Crescimento
Radicular De Feijão Caupi
Sob diferentes Níveis De Salinidade. Acta Botanica Brasilica 26(2):
342-349. Melkasa Agricultural Research Center (MARC) (2017).
Melkasa Agricultural Research Center Profile, Melkasa,
Ethiopia.
Minagawa J, Takahashi Y (2004). Structure,
Function and Assembly of Photosystem II and Its Light-Harvesting Proteins.
Photosynthesis Research 82(3):241-263.
Moniruzzaman M, Islam MN, Hossain MFB, Rashid MM, Ahamed KU (2013). Evaluation of Tomato Germplasm against Salinity. Bulletin of the
Institute of Tropical Agriculture, Kyushu University 36:9-16.
Munns R, Termaat A (1986).
Whole-Plant Responses to Salinity. Functional Plant
Biology 13(1):143-160.
Munns R (2002). Comparative Physiology of Salt and Water Stress. Plant, Cell
and Environment 25(2):239-250.
Munns R, Hare RA, James RA, Rebetzke GJ (2000). Genetic Variation for
Improving the Salt Tolerance of Durum Wheat. Australian Journal of
Agricultural Research 51(1): 69-74.
Murata N, Takahashi S, Nishiyama Y, Allakhverdiev SI
(2007). Photoinhibition of Photosystem II under Environmental Stress. Biochimica et Biophysica
Acta (BBA)-Bioenergetics 1767(6):414-421.
Nishiyama Y, Allakhverdiev
SI, Murata N (2011). Protein Synthesis is the Primary Target of Reactive Oxygen
Species in the Photo inhibition Of Photosystem II. Physiologia
Plantarum 142(1):35-46.
Olumana M, Lois K, Furst J
(2009). Effect of Lake Basaka
expansion on the sustainability of Matahara SE in the
Awash River basin, Ethiopia. Water, Sanitation and Hygiene: Sustainable
Development and Multi-sectorial Approaches. 34th
WEDC International Conference, Addis Ababa, Ethiopia, 2009.
Pak VA, Nabipour M, Meskarbashee M
(2009). Effect of Salt Stress on Chlorophyll Content, Fluorescence, Na+
and K+ Ions Content in Rape Plants (Brassica napus L.). Asian Journal
of Agricultural Research
3(2):28-37.
Raghavendra AS (1991). Physiology of Stress.
John Wiley and Sons, Inc. United States.
Rahneshan Z, Nasibi F, Moghadam AA (2018). Effects of Salinity Stress on Some
Growth, Physiological, Biochemical Parameters and Nutrients in Two Pistachio (Pistacia vera
L.) Rootstocks. Journal of Plant Interactions
13(1):73-82.
Ripoll J, Urban L, Staudt M, Lopez-Lauri F, Bidel LPR, Bertin N (2014). Water
Shortage and Quality of Fleshy Fruits-Making the Most of the Unavoidable,
Journal of Experimental Botany 65(15):4097–4117.
Rozema J, Cornelisse D,
Zhang Y, Li H, Bruning B, Katschnig
D, Broekman R, Ji B, van Bodegom P (2015). Comparing Salt
Tolerance of Beet Cultivars and their Halophytic Ancestor: Consequences of
Domestication and Breeding Programmes. AoB
Plants P. 7.
Saleem A, Ashraf M, Akram
NA (2011). Salt (NaCl) ‐ Induced Modulation in Some Key Physio‐Biochemical Attributes in Okra (Abelmoschus esculentus
L.). Journal of Agronomy and Crop Science 197(3):202-213.
Satoh K,
Smith CM, Fork DC (1983).
Effects of Salinity on Primary Processes of Photosynthesis in
the Red alga Porphyra perforata.
Plant Physiology 73(3):643-647.
Sholi NJ (2012). Effect of Salt
Stress on Seed Germination, Plant Growth, Photosynthesis and Ion Accumulation
of Four Tomato Cultivars. American Journal of Plant Physiology
7:269-275.
Sousa de, JR, Gheyi HR, Brito ME, Lacerda CFD, Silva FVD, Soares FA
(2016). Quantum Efficiency of Photosystem II and Production
of Orange under Salt Stress and Nitrogen Fertilization. Revista Brasileira de Engenharia Agrícola e Ambiental 20(5):434-440.
Sta-Baba R, Hachicha
M, Mansour M, Nahdi H, Ben Kheder
M (2010). Response of Onion to Salinity. The African
Journal of Plant Science and Biotechnology 4(2):7-12.
Stanisavljevic NS, Nikolic DB, Jovanovic ZS, Samardzic JT, Radovic SR,
Maksimović VR (2011). Antioxidative Enzymes in the
Response of Buckwheat (Fagopyrum esculentum Moench) to Complete Submergence. Archives of
Biological Sciences 63(2):399-405.
Stavridou E, Hastings A, Webster RJ, Robson PR (2017). The Impact of Soil Salinity on the Yield,
Composition and Physiology of the Bio-Energy Grass
Miscanthus× giganteus. Gcb Bioenergy
9(1):92-104.
Wani AS, Ahmad A, Hayat S, Fariduddin Q (2013). Salt-Induced Modulation in Growth,
Photosynthesis and Antioxidant System in Two Varieties of Brassica juncea. Saudi Journal of
Biological Sciences 20(2):183-193.
Zelleke A, Derso E (2015).
Production and Management of Major Vegetable Crops in Ethiopia. 32-51. Ethiopian Institute of Agricultural Research. Addis
Ababa, Ethiopia.
