Among abiotic stresses, drought is the most severe abiotic stress in many
parts of the world and is one of the grand problem in present-day climatic
scenario. Drought is the prominent environmental stress, which is
significantly hampering crop yield and its quality in the world. Climate
change increases the odds of worsening drought in many parts of the world in
the decades ahead, it damages the crop that has occurred because of abnormal
metabolism and may reduce growth and death of crop development. The
physiological activity of the crop is also influenced by water stress
through suppressing photosynthesis and the consumption of assimilates in the
expanding leaves. Crop production is determined by the existence of
sufficient rain fall, especially in areas where crop production totally rely
on rain fall, there is always risk of crop failure or yield loss due to
moisture stress. In severe cases, moisture stress could lead to total crop
loss. Drought stress occurs at different stages of growth and adversely
affect yield and yield related traits, which leads to reduction in yield.
The effect of drought stress is mainly depending on the developmental stage
of the plant, degree and duration of the stress, genotypic capacity of
species and environmental interactions. Crop plants have adaptation
strategies to survive under drought stress by the development of various
morphological, physiological and biochemical mechanisms. However, a plant
may exhibit more than one strategy to cope with drought stress. Drought
resistance is the mechanism(s) causing minimum loss of yield in a drought
condition. Drought escape, dehydration avoidance, reduced transpiration or
physiological factors are some drought resistance mechanisms. Eventually,
global food security is threatened by climate change and the most
challenging in the 21st century, affecting the supply of sufficient food for
the increasing world population. The use of well-adapted and high-yielding
varieties with resistance to drought stress is important to reach maximum
yield potential as long as possible through minimizing the risk of climate
change. Climate-smart agriculture is the only way to reduce the negative
impact of climate variations on crop adaptation, before it might affect
global crop production drastically.
The growing global population coupled with the increasing
challenges of climate change, limited arable land and environmental stressors
pose significant threats to global food security. Meeting the ever-rising
demand for food requires innovative approaches to enhance crop productivity
while minimizing the environmental impact of agriculture climate (Lesket al.,
2016). Global food security is being haunted by the rapid increase in
population and drastic changes in the climate (Lesket al., 2016). In the wake of changing
climate, drought and heat stress have become the most important limiting
factors to crop productivity and ultimately the food security. The reduced
precipitation and changed rainfall patterns are causing the frequent onset of
droughts around the world (Lobellet al., 2011). Severe droughts cause substantial
decline in crop yields through negative impacts on plant growth, physiology and
reproduction (Barnabas et al., 2008).
Plants are subjected to the drought
conditions when either the water supply to the roots is limited or the loss of
water through transpiration is very high (Anjumet al., 2011). The severity of the
damage caused by the drought is generally unpredictable as it is driven by
various factors including the rainfall patterns, moisture-holding capacity of
the soil, and water losses through evapotranspiration. Drought interferes with
growth, nutrient and water relations, photosynthesis, assimilate partitioning
and ultimately cause a significant reduction in crop yields (Farooqet al.,
2009). The plant response to drought stress generally varies from species to
species depending on plant growth stage and other environmental factors (Demirevskaet al.,
2009).
Drought is one of the major
environment constraints that limits agricultural production worldwide and leads
to the lack of adequate moisture that is required for normal plant growth and
development and to complete their life cycle (Chen et al., 2020). Drought stress severely affects the plants by
causing substantial reductions in the crop growth and biomass accumulation. The
main consequences of drought stress in plants are the reduced rate of cell
division and expansion, root proliferation, stem elongation and leaf size.
Drought also disturbs the stomatal oscillations,
plant water and nutrient relations that result in declining the crop
productivity, and water use efficiency (Anjumet al., 2011). Drought is a significant limiting factor for
agriculture, and it is the leading cause of crop yield reduction. The
identification of genetic factors involved in plant responses to drought stress
will pave the way for breeding drought-resistant plants.
Food productivity is decreasing due to
detrimental effects of various biotic and abiotic stresses; therefore
minimizing these losses is a major area of concern to ensure food security
under changing climate. Environmental abiotic stresses, such as drought,
extreme temperature, cold, heavy metals or high salinity severely impair plant
growth and productivity worldwide. Drought being the most important
environmental stress, severely impairs plant growth and development, limits
plant production and the performance of crop plants, more than any other
environmental factor (Shao et al.,
2009). Plant experiences drought stress either when the water supply to roots
becomes difficult or when the transpiration rate becomes very high. Drought
impacts include growth, yield, membrane integrity, pigment content, osmotic
adjustment water relations, and photosynthetic activity (Prabaet al., 2009). Drought stress is
affected by climatic, edaphic and agronomic factors. The susceptibility of
plants to drought stress varies in dependence of stress degree, different
accompanying stress factors, plant species and their developmental stages (Demirevskaet al.,
2009).
This day, there are multiple production
constraints, particularly rapid population growth and climate changes are two
critical issues that require immediate action to achieve sustainable
development goals. The rising population is posing increased demand for food,
thereby pushing for an acceleration in agricultural production. Therefore,
increasing crop yield is required to meet the needs of increasing population
growth, however yield reduction is observed in areas where drought is
predominantly devastated crop production. Short duration drought stress mostly
reduces grain yield while prolonged drought stress leads to complete death of
plant. Improving the crop traits is highly required for the development of
superior crop varieties to deal with climate change and the associated abiotic
and biotic stress challenges. Climate change-driven global warming can trigger
higher insect pest pressures and plant diseases thus affecting crop production
sternly. The traits controlling genes for stress or disease tolerance are
economically imperative in crop plants.
Generally, drought is a periodic phenomenon
that endangers crop yields and threatens the livelihoods of populations all
over the world (Liedtkeet al., 2020). The development of drought resistant crop varieties
through breeding or biotechnology is a major challenge for agriculture.
Understanding how drought affects plants is therefore crucial for designing
superior cultivars with consistent high yields. Plant responses to drought
stress, on the other hand, are complicated and vary based on environmental
conditions, stress frequency and duration, plant species and variety, and
physiological stage at the time of stress. Drought limits the agricultural
production by preventing the crop plants from expressing their full genetic
potential. Global climate change gives rise to
numerous environmental causes including biotic and abiotic stresses, which
affect crop productivity (Razaet al., 2023).
Among them, drought stress is a destructive natural threat to food security,
affecting a substantial fraction of the overall population, mainly those living
in arid and semi-arid areas (Raiet al., 2021).
Drought
stress can usually be described as a prolonged time of irregular,
lower-than-average natural water accessibility because drought stress mainly
occurs from a substantial shortage in humidity supply as precipitation.
Generally, plants are exposed to the drought stress when (a) the water transfer
to the roots is inadequate or (b) the water loss via transpiration is extremely
high (Cheng et al., 2021). The damage resulting from drought stress
severity is usually unstable, as several aspects such as the rainfall forms
determine it, moisture availability in soil, and water deficiencies because of
transpiration. Consequently, drought stress hinders crop growth, water-nutrient
relations, and photosynthesis and eventually triggers a substantial decline in
crop yields (Mubariket al., 2021). Plant responses to drought stress usually
differ from species to species, varying on growth phase and further environmental
influences (Ansari et al., 2019).
Drought
stress is a critical agronomic issue that causes severe production losses
around the world. Developing crops that are well adapted to drought-prone
environments could help to alleviate this agricultural constraint. The
mechanisms that allow this crop to thrive in such severe environments are
complicated and poorly understood. Drought in agriculture, specifically water
scarcity, has a negative impact on plant and crop productivity by reducing leaf
size, stem extension, and root proliferation, disrupting plant water and
nutrient relationships, and diminishing water-use efficiency. During periods of
severe drought, these losses can be significantly larger, and crop failure is a
distinct possibility. Drought is a major constraint in sorghum production
around the world, and it is the leading cause of crop yield reduction (Sabadinet al.,
2012).
To
cope with drought stress, plants reprogram a wide range of responses at the
molecular, biochemical, and physiological levels (Thatcher et al., 2016). Depending on the tissue type, developmental stage,
and stress level, these changes can happen rapidly and with a lot of precision.
At the molecular level, drought stress causes transcriptional and
post-transcriptional regulation of gene expression (Takahashi et al., 2018). Differential expression
of genes involved in various metabolic pathways is caused by transcriptional
modulations, resulting in changes in metabolite flow and physiological changes
associated to cellular damage protection (Knight H and Knight M.R, 2001).
In
the process of evolution, plants have developed complex regulatory networks to
cope with drought, including drought escape, drought avoidance, drought
tolerance, and drought recovery after stress. Three mechanisms, namely, drought
escape, drought avoidance and drought tolerance are involved in drought
resistance. Various morphological, physiological and biochemical characters
confer drought resistance. Morphological and physiological characters show
different types of inheritance pattern (monogenic or polygenic) and gene action
(additive and non-additive), whereas the genes responsible for biosynthesis of
different compatible solutes have been identified and cloned from plants,
yeasts, mouse and human. Different breeding approaches for drought resistance have
emerged with their merits and demerits. Efficient screening techniques are
pre-requisite for success in selecting desirable genotypes through any breeding
program. The objective of the paper was to understand the impact of drought
stress and genetic drought resistance mechanism under water-limited
environments.
2. DROUGHT RESISTANCE MECHANISMS IN CROP
PLANTS
Plants are influenced by both biotic and
abiotic factors, and in response to these factors, numerous internal changes
occur in plants. These biotic and abiotic factors influence plant growth and
development along with productivity. Biotic factors are interactions of
organisms with plants that have both positive and negative effects. Positive
effects may have a beneficial influence on plant growth. Negative effects may
include allelopathy, herbivory
influence, or pathogen infection in plants (Ciura and
Kruk, 2018). Plant defense systems with various chemical
compounds help to resist those negative effects (Li et al., 2019).
Drought
resistance is mechanisms causing minimum loss of yield in a drought
environment. Different mechanisms through which a crop is capable of minimize
the loss in yield due to drought stress. In response to drought stress, plants
activate their drought response mechanisms, such as morphological and
structural changes, expression of drought-resistant genes, synthesis of
hormones, and osmotic regulatory substances to alleviate drought stress. Drought resistance or tolerance is a
broader term applied to plant species with adaptive features that enable them
to escape, avoid, or tolerate drought stress (Levitt, 1980).
Critical
evaluation of progress in plant breeding over a period of several decades has
demonstrated a genetic improvement in yield under both favorable and stress
conditions (Castleberry, Crum & Krull, 1984). The
yield improvement under drought stress occurred before many of the
physiological issues of drought resistance were understood and resulted partly
from the genetic improvement of yield potential and partly from the improvement
of stress resistance. For example, Bidingeret
al. (1987) found that the yield of millet varieties under drought
stress was largely explained by their yield potential and growth duration.
Early varieties with a high yield potential were most likely to yield best
under stress. Fischer & Maurer (1978) also recognized the effect of
potential yield on yield performance of wheat under drought stress and proposed
a ‘susceptibility index’ (S) which estimated the relative susceptibility of a
variety to drought stress. In analyzing their wheat data, they found that
susceptibility index was not very independent of the potential yield of the
variety.
The
improvement of yield under stress must therefore combine a reasonably high
yield potential (Blum et al., 1983)
with specific plant factors, which would buffer yield against a severe
reduction under stress. On the other hand, potentially lower yielding genotypes
occasionally have been found to perform very well under drought stress
conditions especially under severe drought stress (Blum, 1982). One is left
with the long-standing practical conclusion of Reitz (1974) that ‘Varieties
fall into three categories: (a) those with uniform superiority over all
environments; (b) those relatively better in poor environments; and (c) those
relatively better in favored environments’.
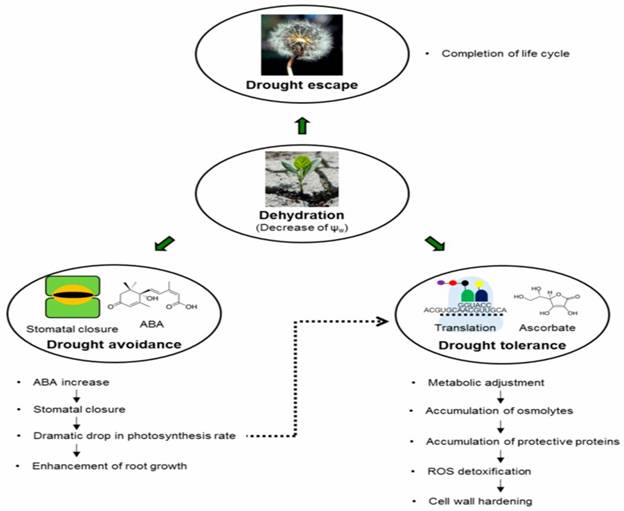
Figure
1: The
main drought resistance strategies employed by plants to counter water deficit
periods (drought escape, drought avoidance, and drought tolerance) and the main
steps of the plant response to dehydration.
There are four different mechanisms, which help in survival of plants
under moisture deficit conditions. There are drought
recovery, drought avoidance, drought tolerance and drought escape.
2.1 Drought Escape
The simplest
way of survival under drought conditions is to escape drought. Generally,
drought occurs either in the mid or late-crop season. Drought escape is most
common in case of plants grown in desert regions. They complete their life
cycles in 4 to 6 weeks. Drought escape also plays an important role in some crop
plants. For Example, yields of early varieties of wheat, sorghum, maize, and
rice arc less affected by severe drought than late maturing ones. All these
crops have determinate growth habit. In spring wheat, late maturing varieties
give higher yield than early types especially when drought occurs early in the
season and is over before anthesis.
2.2 Drought Tolerance
The ability of crop plants to withstand low tissue water
content is referred to as drought tolerance. Drought tolerance is more
desirable because the crop can produce more yield at lower water potential. In
cereals, drought tolerance generally operates during reproductive phase.
Tolerant cultivars exhibit better germination, seedling growth and
photosynthesis. In Sorghum, a drought resistant line exhibited higher
photosynthetic rate at leaf water potential than a less drought resistant line.
Drought tolerance differs from drought avoidance in several aspects.
Table-1: Traits Associated With Drought
Tolerance
No
Category
Traits
1
Morphological & Anatomical
Yield; More Root length, Root Volume, Root Dry Weight, Root Thickness;
Root surface area, More Plant Biomass; Harvest index; Leaf drying; Leaf tip
firing; Delay in flowering.
2
Phenological
Early to maturity, Late Flowering; Anthesis,
Silking Interval; Seedling vigor; Weed
competitiveness; Photosensitivity; perennially.
3
Physiological & Biochemical
Osmotic Adjustment; Carbon Isotope Discrimination; Stomatal
conductance; Remobilization of stem reserves; Specific leaf weight; ABA;
Electrolyte leakage; leaf rolling, tip firing, Stay-green; Epicuticular wax; Feed forward response to stress; Heat
shock proteins; Cell wall proteins; Leaf water potential; Water use efficiency;
Aquaporins; Nitrogen use efficiency; Dehydrins.
2.3 Drought Avoidance
Drought avoidance refers to
ability of the plant to maintain a favorable internal water balance under
moisture stress. In other words, plants that avoid drought retain high water
contents in their tissues. Drought avoidance can permit a longer growth period
in the crop through reduced water use or increased water uptake. However,
drought avoidance leads to reduction in photosynthesis and thereby reduction in
the growth of aerial parts. It leads to increase in root development and
therefore, is more important than drought tolerance. In cereals, drought
avoidance operates during vegetative phase, while tolerance operates during,
reproductive phase. Drought avoidance mechanisms are of two types. First, those
that reduce water loss through transpiration. Such features include stomatal characteristics and shape, size and orientation of
leaves. The second, those that maintain water uptake during drought period.
2.4 Drought Resistance
Drought resistance is the sum of drought
avoidance and drought tolerance. In other words, drought resistance refers to
the ability of crop plants to give good yield under moisture deficit
conditions. Drought resistance is measured in terms of various mechanisms
associated’; with drought tolerance and yield under soil moisture deficit. In
winter wheat, both avoidance and tolerance features are important for drought
resistance.
3. MAJOR
TRAITS CONTRIBUTING TO DROUGHT RESISTANCE
A range of morphological and physiological traits have
been linked to drought tolerance in plants, which include root morphology and
rooting depth, plant architecture, leaf area, cuticular
resistance and thickness, stomatal conductance,
osmotic adjustment, antioxidative defense, hormonal
regulation, desiccation. The most important include root architecture, leaf
morphology, physiological characters such as osmotic adjustment or proline accumulation, partitioning of total biomass
(determined by dry matter or harvest index), timing of plant development (e.g.
earliness), or others associated with the plant reproductive biology. Some of
these characteristics are specific while others are common for many species.
Some reports indicate a significant association between crop tolerance to heat
and respective adaptation to drought-prone environments in the warm tropics.
3.1 Leaf Traits: Senescence, Stay-Green, and Leaf Area
Plant functional traits are useful tools for
exploring how plants adapt to the environment and studying global climate
change (Fyllaset al., 2020). Among these traits, leaf traits have received
particular attention due to their sensitivity to climate change and their
ability to reflect plant resource acquisition and utilization (Ye et al., 2022). In dry conditions, plants tend to have thicker leaf
thickness (LT), higher leaf dry mass per area (LMA), and larger leaf dry matter
content (LDMC), in order to reduce water loss and enhance their ability to
adapt to the drought environments (Akramet al., 2022). Leaf nitrogen content is closely related to
photosynthesis (Zhan et al., 2018). The leaf carbon capture strategy can be represented by
nitrogen content per unit area (Narea), nitrogen
content per unit mass (Nmass), and carbon: nitrogen
ratio (C/N) (Zhan et al., 2018). Plants typically had higher nitrogen content per unit
area and higher leaf dry mass per area under hot and dry environmental
conditions, as this increased investment of nitrogen in structure enhanced
their survival in adversity (Blumenthal et al., 2020). As essential members of plant functional traits, leaf
traits can provide insight into the relationship between plants and the
environment at both the regional and global scales (Toledo-Aceveset al.,
2022).
Senescence is a developmental stage of plant
leaves that leads to the arrest of photosynthesis, the degradation of
chloroplasts and proteins, and the mobilization of nitrogen, carbon, and other
nutrient resources from the leaves to other organs. As most cereals are
monocarpic annual species, these resources are directed to developing seeds,
and senescence therefore plays a relevant role in crop yield. Environmental
stresses like temperature, lack of nutrients, and drought might initiate
senescence prematurely, affecting seed nutritional composition and crop yield (Distelfeldet al., 2014). In crops
threatened by terminal drought, the ability to sustain photosynthetic activity
longer by delaying or slowing down senescence could be an effective strategy to
avoid yield losses. Plant
breeders commonly refer to the trait that confers extended photosynthetic
activity as stay-green, also defined as green leaf area at maturity (GLAM).
This trait is well studied in sorghum, a dry climate-adapted cereal in which a
number of stay-green quantitative trait loci (QTLs) have been identified (Vadezet al., 2011). However, the genes underlying these QTLs have not yet
been identified (Harris-Shultz et al., 2019).
Stay-greenness
in sorghum is a complex trait, and it is connected with the perennial
tendencies of some varieties (Thomas and Howarth, 2000). Other
plant species achieve stay-green characteristics via substantially
different pathways that include disabling chlorophyll catabolism (like in the
case of Gregor Mendel’s green peas), (Armsteadet al., 2007), and altering the responses to plant hormones. Indeed,
some stay-green genes have also been identified in Arabidopsis and rice (Hortensteiner,
2009), notably the stay-green rice (SGR)
genes and their homologs in Arabidopsis SGR1, SGR2, and SGR-like
(SGRL). The respective molecular pathways have been elucidated, with the phytohormones ethylene, ABA, cytokinin
(CK), and strigolactone (SL) having a prominent role
in stress-induced leaf senescence (Abdelrahmanet al., 2017). The connection between ethylene and leaf senescence is
long known (Bleeckeret al., 1988), and numerous attempts to improve photosynthetic
activity and drought performance by manipulating ethylene biosynthesis have
been published in dicots (John et al., 1995) and cereal plants (Young et al., 2004).
3.3 Stomatal-Mediated
Drought Responses
Stomata, which are openings on the surface of
the aerial portion of plants, are enclosed by two specialized guard cells that
can open and close the pore by changing their turgor pressure. Stomata are
vital for CO2 uptake in photosynthetic organs and are finely
regulated by a molecular pathway that allows plants to acquire CO2 while
minimizing water loss. Manipulating stomatal number,
size, and regulation was one of the earliest strategies adopted by scientists
in attempt to produce drought-resistant plants, and recent advances
in Arabidopsis and crops. The main hormone signal that triggers stomatal closure in water-limited conditions is ABA (Sussmilch and
McAdam, 2017).
The manipulation of ABA sensitivity to increase stomatal
responses in response to drought could help plants to survive. However,
diminished photosynthetic activity due to limited CO2 uptake is
usually detrimental to carbon assimilation and negatively affects crop yield.
In addition, water evaporation through stomatal
openings prevents plants from overheating.
Roots are among the first defense towards drought with other morpho-physiological and biochemical mechanisms employed by
plants. The crop root system (Hulugalleet al., 2015) plays an
important role in water and nutrient uptake, which largely determines the
environmental adaptability and yield of crops. Because of the difficulty of
underground research, the research of the plant root system is far behind that
of plant aboveground part. For crops, the main strategy to cope with drought is
to regulate root growth and root structure. Some studies have assessed breeding
and field management, and considered that they have the same significance in
improving crop yield (Li et al., 2017).